Research Report
Taxonomic Revision of the Tiger Fish Hydrocynus vittatus (Castelnau, 1861), H. Brevis (Cuvier & Valencience, 1849) and H. forskalii (Cuvier, 1819) from the Nile in Sudan 
 Author
Author  Correspondence author
Correspondence author
International Journal of Aquaculture, 2015, Vol. 5, No. 5 doi: 10.5376/ija.2015.05.0005
Received: 06 Feb., 2015 Accepted: 08 Mar., 2015 Published: 06 May, 2015
Mohamed and Al-Awadi, 2015, Taxonomic Revision of the Tiger Fish Hydrocynus Vittatus (Castelnau, 1861), H. Brevis (Cuvier & Valencience, 1849) and H. Forskalii (Cuvier, 1819) from the Nile in Sudan, International Journal of Aquaculture, Vol.5, No.5: 1-9 (doi: 10.5376/ija.2015.05.0005)
The main objective of this study was to revise the taxonomic status of three species from the genus Hydrocynus: H. vittatus (Castelnau, 1861), H. brevis (Cuvier & Valencience, 1849) and H. forskalii (Cuvier, 1819), commonly known as "Tigerfish" to determine the morphological characters for distinguishing them. The study was based on morphometric and meristic characters plus body weight of specimens. Analysis of morphological characters was done by the statistical programme “PAST”. The study also considered the length – weight relationships and the type of growth for each species. Twenty morphometric characters had almost equal loads, ranging between (-0.1843 and -0.2436) in separating the three species. The number of lateral line scales, LLS, had the biggest load of (-0.9994) followed by the number of teeth in upper jaw, UJT (-0.5643), the pectoral fin rays, PFR (-0.4638), gill rakers, GR (-0.4596) and the number of teeth in lower jaw, LJT (0.3471). The morphological results indicated that a combination of morphological characters can be used to separate the three species of the genus Hydrocynus. They may also be distinguished by the position of the dorsal fin compared to insertion of the ventral; the black-edge of the dorsal and adipose fins and the black-edge fork caudal fin of H. vittatus; and the reddish-orange lower caudal-fin lobe and the red anterior area of anal fin of H. brevis. The length-weight relationship was W = 3.088 L-2.079 for H. vittatus and (r2 = 0.923); W = 3.451 L-2.534 for H. brevis and (r2 = 0.664); W = 2.9981L-1.9815 and (r2 = 0.418) for H. forskalii. The values (b = -2.079, -2.534 and --1.9815) indicated allometric growth of the three species. There was a strong positive correlation between standard body length and body weight of H. vittatus. It is recommended that morphological analysis should be compared and combined with biochemical and molecular phylogenetics.
Hydrocynus is a genus of large characin fish, in the family Alestidae that contains five species (Boulenger, 1907), with three species common in the Nile (Sandon, 1950). These are namely H. vittatus (Castelnau, 1861), H. brevis (Cuvier & Valencience, 1849) and H. forskalii (Cuvier, 1819). The three species share similar common names "Tiger-fish, Kas and Kalb elbahr" by the native fishers in Sudan and share similar distribution in the White Nile, Blue Nile and the Nile to Lake Nubia, but H. forskali is the commonest species (Bailey, 1994). Like H. vittatus, H. brevis is deep bodied with dark spot on the adipose fin (Bailey, 1994). Hydrocynus brevis is easily distinguished from H. forskalii by shorter body covered with smaller scales and small eyes, but the two species are not distinguished under special names by the native fishermen (Sandon, 1950). The synonymy of H. forskalii with H. vittatus was revised by Brewster (1986). However, all of them are rather similar in appearance, their distinction requiring careful examination.
Therefore, the present study was designed to revise and compare the morphological characteristics of the three species by using a combination of both morphometric and meristic characters. It is an attempt to taxonomically characterize each species in the Nile waters of Sudan, and determine the morphological characteristics that contribute mostly to the variation of the three species. Analysis based on measurement of morphological characters was done using the Paleontology Statistical program “PAST”. This is a software package originally aimed at Paleontology but currently has become popular in taxonomy, ecology and other fields (Hammer et al., 2001). It is a tool highly recommended for determining the relationships between species and populations of a species (Thorpe, 1987; Mwanja et al., 2011; Mohamed, 2014; Mohamed and Awad Elseed, 2014). The study also considered the length – weight relationships and the type of growth for each species. A clear description of each species is given here after.
2 Material and Methods
2.1 Collection of fish
Specimens of Hydrocynus species were purchased from the Central Market in Khartoum. Comparative material included 30 specimens (10 from each of H. vittatus (Castelnau, 1861), H. brevis (Cuvier & Valencience, 1849) and H. forskalii (Cuvier, 1819). Each individual specimen was identified according to the original description of Boulenger (1907). The institutional abbreviations followed Daget and Grosse (1984).
2.2 Morphometric measurements
Twenty-eight morphometric measurements and ten meristic counts were taken for each specimen according to Teugels and Thys (1990). All measurements were taken on the right side of the specimens and were point-to-point measurements taken by a fine dial caliper to (0.00) mm. For descriptive purposes all measurements were expressed as ratios of standard length (%SL). The measurements of head structures and inter-orbital width were expressed as percentage of head length (%HL). Principal component analysis (PCA) was used to explore the multivariate variable data matrix to reduce the large number of variables into a few biologically meaningful axes (principal components) that explain as much variations as possible (Past, 2005). Raw data of morphometric measurements (not meristic counts) was transformed to logs10 and used for multivariate analysis. Morphometric measurements were log10- transformed to correct for length differences. The loadings of the variables were done to determine their importance on variability explained. Cluster analysis of morphometric and meristic characters was performed separately to identify the similarity of individuals of each subspecies. Morphometric measurements and meristic counts were analyzed separately. The length–weight relationship and type of growth of pooled data for each species were estimated by the equation W = aLb, where W = weight of fish in grams, L = SL in cm, b = length exponent (slope) and a = proportionality constant (intercept) according to (Bagenal, 1978). The correlation or degree of association (r2) between length and weight was calculated from linear regression analysis.
3 Results and Discussion
Twenty morphometric characters expressed as percentage of standard length, % SL, and three morphometric characters expressed as percentage of head length, % HL, plus head length, were found to be significantly different (P < 0.05) between the three species (Table 1 & 2). The three species differ significantly (p < 0.05) in the number of rays of pectoral and anal fins, where H. vittatus has more fin rays compared to the other two species (Table 3). More scales of the lateral line; LLS; were recorded in H. brevis, and Dorsal– to– lateral line scales, DLS; gill rakers, GR and teeth in the lower jaw, LJT are significantly higher in H. vittatus.
.jpg) Table 1 Morphomertic measurements of H. vittatus (A), H. brevis (B) and H. forskalii (C) and expressed as percentage of standard length (%SL) |
.jpg) Table 2 Morphomertic measurements H. vittatus (A), H. brevis (B) and H. forskalii (C) expressed as percentage of head length (%HL) |
.jpg)
Principal component analysis of data from the 27 morphometric measurement revealed that approximately 70.3% of the total variation was explained along one component, (Table 4). The second component of variation accounted for 11.5% of the total variability, the third component of variation accounted for 4% of the total variability and the fourth component of variation accounted for 3% of the total variability. The Eigenvalues for all components were positive indicating that all used variables has some effect on the morphological variation of the Hydrocynus species The loadings of the morphometric variables to determine their importance on variability explained is presented in (Figure 1). Principal component analysis of data from ten meristic counts revealed that approximately 73.9% of the total variation was explained along one component (Table 5). The second component of variation accounted for 11.6% of the total variability, the third component of variation accounted for 4.5%, the forth component of variation accounted for 3.3% and the fifth component of variation accounted for 3.1% of the total variability. The loadings of the meristic variables to determine their importance on variability explained is presented in (Figure 2).The data of log10–transformation of morphometric measurements and cluster analysis of meristic counts produced hierarchical clusters of specimens of the three species in a distance dendrogram. Most individuals of each species clustered together at the end of the spectrum (Figure 3 & 4).
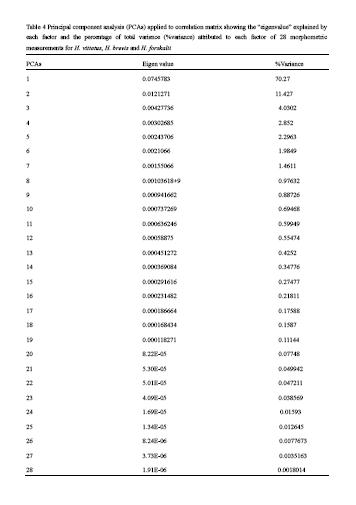 Table 4 Principal component analysis (PCAs) applied to correlation matrix showing the “eigenvalue” explained by each factor and the percentage of total variance (%variance) attributed to each factor of 28 morphometric measurements for H. vittatus, H. brevis and H. forskalii |
Figure 1 A diagram based on morphometric measurements of H. vittatus, H. brevis and H. forskalii specimens, showing the load of each character on their separation |
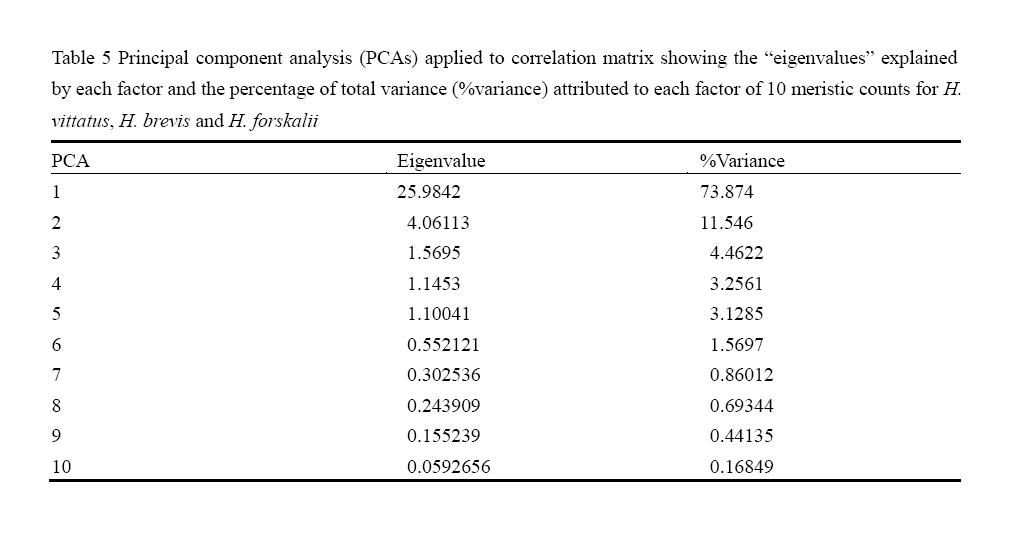 Table 5 Principal component analysis (PCAs) applied to correlation matrix showing the “eigenvalues” explained by each factor and the percentage of total variance (%variance) attributed to each factor of 10 meristic counts for H. vittatus, H. brevis and H. forskalii |
Figure 2 A diagram based on meristic counts of H. vittatus, H. brevis and H. forskalii specimens, showing the load of each character on their separation |
Figure 3 A division hierarchical cluster of log10 transformed of morphometric measurements of of H. vittatus (A), H. brevis (B) and H. forskalii (C), based on a matrix of distance of Neighbour-Joining clustering(nearest neighbor), using Euclidean similarity measure |
Figure 4 A division hierarchical cluster of meristic counts of H. vittatus (A), H. brevis (B) and H. forskalii (C), based on the matrix of distance of Neighbour-Joining clustering(nearest neighbor), using Euclidean |
The length-weight relationship was W = 3.088 L-2.079 equal to (Log10W = 0.48967729 + (-2.0794) Log10SL) for H. vittatus and r2 = 0.923 (Figure 5a); W = 3.451 L-2.534 equal to (Log10W = 0.53795754 + (-2.5335) Log10SL) for H. brevis and r2 = 0.664 (Figure 5b); (Log10W = 0.47684611 + (-1.9815) Log10SL) equal to W = 2.9981 L-1.9815 and r2 = 0.418 for H. forskalii (Figure 5c).
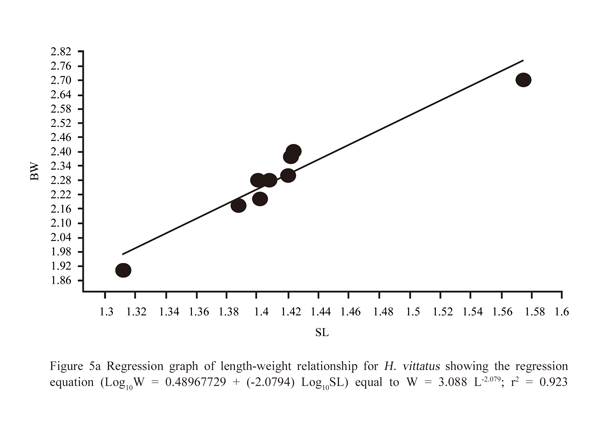 Figure 5a Regression graph of length-weight relationship for H. vittatus showing the regression equation (Log10W = 0.48967729 + (-2.0794) Log10SL) equal to W = 3.088 L-2.079; r2 = 0.923 |
.png) Figure 5b Regression graph of length-weight relationship for H. brevis showing the regression equation (Log10W = 0.53795754 + (-2.5335) Log10SL) equal to W = 3.451 L-2.534; r2 = 0.664 |
 Figure 5c Regression graph of length-weight relationship for H. forskalii showing the regression equation (Log10W = 0.47684611 + (-1.9815) Log10SL) equal to W = 2.9981 L-1.9815; r2 = 0.418 |
Eighteen morphometric characters had almost equal loads, ranging between (-0.1843 and -0.2436) in separ-ating the three species as shown in (Figure 1). Out of the ten meristic characters, the number of lateral line scales, LLS. had the biggest load of (-0.9994) to separate the three species of Hydrocynus, followed by the number of teeth in upper jaw, UJT (-0.5643), the scales between dorsal fin and lateral line, the pectoral fin rays, PFR(-0.4638), gill rakers, GR (-0.4596) and the number of teeth in lower jaw, LJT (0.3471) as shown in (Figure 2). Although all these characters have negative loads but can be used to separate the two species. Accoding to Snoke (2004), both positive and negative loads can be used to distinguish species.
Cluster analysis of meristic counts from H. vittatus showed less divisions compared to H. brevis and H. forskalii, indicating less variation in meristic counts within this species. Cluster analysis also indicated the existence of some significant morphological differences between the three species, but meristic characters have more significant effect than morphometric ones in separating the three species. The present study confirmed the observations of Bailey (1994) that H. brevis has the deepest body compared to H. vittatus and H. forskalii, but opposite to Boulenger (1909) and Sandon (1950), who reported H. forskalii to have the deepest body.
Boulenger (1909) and Sandon (1950) stated that the position of the dorsal fin was below or slightly behind the ventral in H. vittatus; directly below the ventral in H. brevis and in advance the ventral in H. forskalii. (Paugy et al., 2003) recoded the origin of dorsal-fin at same level as, or slightly before the ventral-fin insertions for H. vittatus; dorsal-fin origin at the same level as, or slightly ahead the level of pelvic-fin insertions in H. brevis and distinctly before the level of pelvic-fin insertions in H. forskalii. In the present study the position of the dorsal fin was found to be distinctly before the insertion of ventral fin in H. vittatus, slightly behind the insertion of ventral fin in H. brevis and directly below or slightly behind the insertion of ventral fin in H. forskalii. The head width/ head length ratio of H. vittatus was smaller compared to the ratios of the other two species, but the CPL/CPD ratio of the three species was equal. Eye diameter was about 77% of interorbital width in H. brevis and H. forskalii, but at least 80% of inter-orbital width in H. vittatus. (Paugy et al., 2003) recorded eye diameter less than 60% of interorbital width in H. brevis, and at least 70% in H. vittatus and H. forskalii. In the present study H. vittatus was found to have (47 - 58) scales in the lateral line (LLS), H. brevis (51 - 65) and H. forskalii (48 - 62). Boulenger (1909) and Sandon (1950) recorded (44 - 48), (47 - 54) and (48 - 54) scales in lateral line of the three species, respectively, and (Paugy et al., 2003) recorded 43-53 lateral line scales for H. vittatus, 47-55 for H. brevis and 47-54 for H. forskalii.
The values of the regression coefficients (b = -2.0794, -2.5335 and -1.9815) indicated allometric growth of the three species; that is growth in which each part of the body grows with changing proportions. The high value of regression coefficient for the pooled data of H. vittatus indicated a strong positive correlation between the standard body length and the body weight, where the weight increased with increasing body length. There was a medium correlation in H. brevis and weak in H. forskalii.
The morphological results obtained in the present study indicated that a combination of morphological characters can be used to separate the three species of the genus Hydrocynus. They may also be easily distinguished by the dimensions of the head, where H. vittatus has the longest head (HL) and higher HW/HL compared to the other species. The further distinctive features are the position of the dorsal fin compared to insertion of the ventral; the black-edge of the dorsal and adipose fins; and the black-edge of the forked caudal fin of H. vittatus; the reddish-orange lower edge of the caudal fin and the anterior area of anal fin of H. brevis. Given that the morphological, biochemical and molecular data are only complementary and not in competition with one another (Chang, 2004; Mohamed, 2010), morphological phylogeny analysis should be compared, contrasted and combined with the analysis of the molecular phylogeny, to more clearly defined taxonomic status of the three species H. vittatus, H. brevis and H. forskalii in the Nile waters from different regions in Sudan.
The three species are very similar in size, the maximum length in each case being a little less than 500 mm The distinctive features are given in (Table 6). A detailed description of each species, based on the
present study, is also provided below.
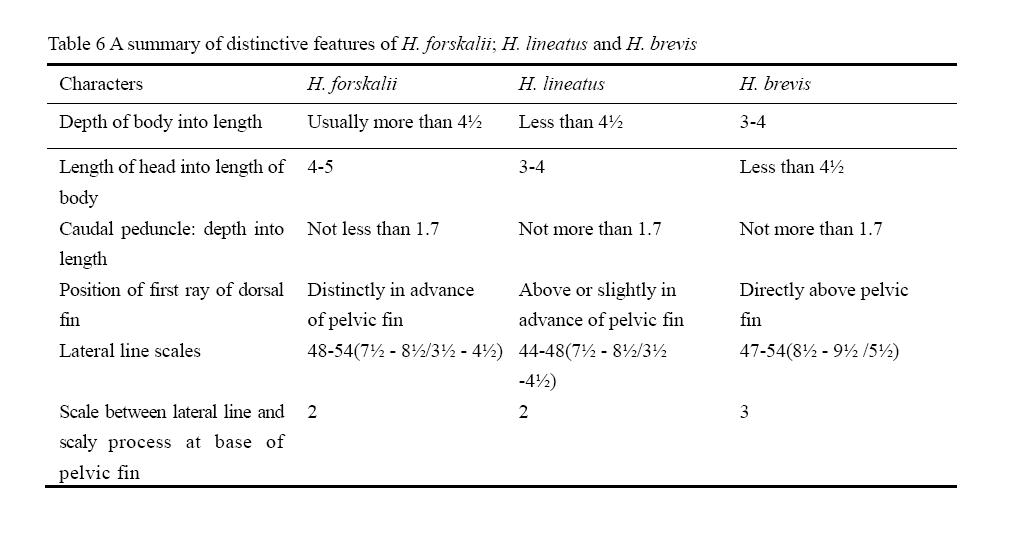 Table 6 A summary of distinctive features of H. forskalii; H. lineatus and H. brevis |
Hydrocynus vittatus (Castelnau, 1861)
Synonyms:
Hydrocinus vittatus Castelnau, 1861
Hydrocyon lineatus Bleeker, 1862
Hydrocyon lineatus Schlegel, 1863
Hydrocyon lineatus Gunther, 1864
Hydrocyon vittatus Boulenger, 1898
Description: A brilliant silvery-coloured fish, with compressed elongated body, 205 to 375 mm standard length, covered by ctenoid scales and each scale marked by a dark spot forming parallel bands visible above the lateral line, and a very small adipose fin behind the dorsal fin. Edges of dorsal and adipose fins black, forked edge of caudal fin distinctly black. Position of the dorsal fin distinctly before the insertion of ventral fin. Eye diameter 80% of inter-orbital width.
Measurements are in %SL: Body Depth 20.4 to 28.9; Head length 23.1 to 27.5; head width 11.2 to 13.1; upper snout length 9 to 11.2; lower snout length 10 to 13.7; eye diameter 6.6 to 9.2; inter-orbital width 9.7 to 11.6; post-orbital length 10.8 to 14.6; dorsal-to- adipose fin 27.7 to 37.5; pre-dorsal length 46.4 to 61.6; pre-ventral length 50.8 to 65.3; caudal peduncle length 10.9 to 12.6; caudal peduncle depth 9 to 11.8.
Measurements are in %HL: Head width 44.7 to 54.1; upper snout length 37.6 to 45.4; lower snout length 40.4 to 55; eye diameter 28.8 to 37.1; inter-orbital width 39 to 46.6; post-orbital length 43.6 to 58.7.
Lateral line scales 47-58; Dorsal fin rays 8 -10; pectoral fin 12 -16; ventral fin 9 - 10; anal fin 12 - 16; gill rakers 11-16; teeth in upper jaw 11-14; teeth in lower jaw 10-14.
Hydrocyon brevis (Günther, 1864)
Synonyms:
Hydrocynus forskalii Cuvier & Valenciennes, 1849
Hydrocynus brevis Günther, 1864
Description: A brilliant silvery-coloured fish, with compressed short body, 185 to 256 mm standard length, covered by ctenoid scales and each scale marked by a dark spot forming parallel bands visible above the lateral line, and a very small adipose fin behind the dorsal fin. Fins of H. brevis are grey, but the lower caudal-fin lobe and the anterior area of anal fin are tinged with reddish-orange. Position of the dorsal fin slightly back the insertion of ventral fin. Eye diameter 77% of inter-orbital width.
Measurements are in %SL: Body Depth 22.5 to 35.3; Head length 24 to 30.6; head width 11.9 to 15; upper snout length 9.8 to 12.8; lower snout length 11.5 to 13.8; eye diameter 8.5 to 10.5; inter-orbital width 10.3 to 13.1; post-orbital length 13.2 to 17; dorsal-to- adipose fin 26.1 to 32.4; pre-dorsal length 49 to 69.4; pre-ventral length 50.7 to 54.5; caudal peduncle length 10.9 to 14.5; caudal peduncle depth 10.5 to 12.6.
Measurements are in %HL: Head width 43.6 to 56.4; upper snout length 36.3 to 43; lower snout length 39.6 to 49; eye diameter 29.2 to 41.5; inter-orbital width 39.3 to 47.7; post-orbital length 54.2 to 61.6.
Lateral line scales 51-65; Dorsal fin rays 10; pectoral fin 13-15; ventral fin 8-9; anal fin 12 - 13; gill rakers 13-15; teeth in upper jaw 12-14; teeth in lower jaw 10-13.
Hydrocynus forskali (Cuvier, 1819)
Synonyms:
Salmo dentexForskal, 1775
Characinus dentex Geoffroy, 1809
Hydrocyon forskalii Cuvier, 1819
Description: General body darker and elongated, 203 to 303 mm standard length, covered by cycloid scales and each scale marked by a dark spot forming parallel bands visible above the lateral line, and a very small adipose. Position of dorsal fin is directly below or slightly back the insertion of ventral fin. Eye diameter 77% of inter-orbital width. Anterior part of anal fin and lower caudal-fin lobe bright red, the other fins uniformly.
Measurements are in %SL: Body Depth 20.2 to 29; Head length 22.2 to 31.2; head width 10.4 to 16; upper snout length 8.9 to 13.8; lower snout length 9.3 to 15; eye diameter 6.8 to 11.5; inter-orbital width 9.8 to 14.7; post-orbital length 13.2 to 19.5; dorsal - to - adipose fin 19.8 to 33.7; pre-dorsal length 43.7 to 68.6; pre-ventral length 39.9 to 64.6; caudal peduncle length 10.8 to 18.6; caudal peduncle depth 9.8 to 15.6.
Measurements are in %HL: Head width 43.9 to 53; upper snout length 35.2 to 56.1; lower snout length 35.7 to 60.1; eye diameter 30.6 to 39.4; inter-orbital width 39.6 to 47; post-orbital length 54.6 to 62.3. Lateral line scales 48 - 62; Dorsal fin rays 9 - 10; pectoral fin 12-14; ventral fin 9; anal fin 12-14; gill rakers 13-15; teeth in upper jaw 12-14; teeth in lower jaw 12.
References
Bagenal T.B., 1978, Methods for Assessment of Fish Production in Freshwater (3rd Edition), Blackwell, Oxford
Bailey R.G., 1994, Guide to the fishes of the River Nile in the Republic of the Sudan, Journal of Natural History, 28: 947-948
http://dx.doi.org/10.1080/00222939400770501
Boulenger G.A., 1907, Zoology of Egypt, The Fishes of the Nile, Hugh Rees Ltd., London
Boulenger G.A., 1909, Catalogue of the Freshwater Fishes of Africa. London. Volume 1
Brewster B., 1986, A review of the genus Hydrocynus Cuvier, 1819 (Teleostei: Characiformes), Bulletin of British Museum of Natural History (Zoology), 50(3): 153-206
Chang M.L., 2004, Phylogenetics analysis of morphological data (Review), Human Biology, 76(1): 165-168.
http://dx.doi.org/10.1353/hub.2004.0017
Daget J., and Gosse J.-P., 1984, Distichodontidae. In: Daget, Gosse, and Thys van den Audenarde (eds), Check-list of Freshwater Fishes of Africa, ORSTOM and MRAC, Tervuren: 184-211
Hammer O., Harper D.A.T., and Ryan P.D., 2001, Paleontological statistics software package for education and data analysis, Paleontology Electronics, 4(1): 4 - 9
Mohamed E.H.A., 2010, Characterization of two Synodontis (Siluriformes: Mochokidae) catfish species in the White Nile and Lake Nubia, Environmental Biology of Fish, 88: 17-23
http://dx.doi.org/10.1007/s10641-010-9585-1
Mohamed E.H.A., 2014, Morphological revision of Ichthyborus besse besse and Ichthyborus besse congolensis (Pisces: Distichodontidae), International Journal of Aquatic Science, 5(2): 181-197
Mohamed E.H.A, and Awad Elseed E.A.G., 2014, Morphological revision of the Nile catfish Bagrus bayad (Forsskal, 1775) and Bagrus docmac (Forsskal, 1775) (Pisces: Bagridae), Journal of Aquatic Biology and Fisheries, 2(1): 105-114
Mwanja M.T., Muwanika V., Nyakaana S., Masembe C., Mbabazi D., Justus R. J., and Mwanja W. W, 2011, Population morphological variation of the Nile perch (Lates niloticus, L. 1758), of East African Lakes and their associated waters, African Journal of Environmental Science and Technology, 5(11): 941-949
Paugy D., Leveque C., and Teugels G.G., 2003, The Fresh and Brackish Water Fishes of West Africa. IRD Editions, Scientific Publications of the Royal Museum for Central Africa, Belgium., 1: 283 - 287
Past, 2005, Statistical Package for Scientific analysis
Sandon H., 1950, An illustrated guide to the freshwater fishes of the Sudan. Sudan Notes and Records, Special Publication, London, McCorquogale: 33-34
Snoeks J., 2004, The Cichlid Diversity of Lake Malawi/Nyasa/Niassa: Identification, Distribution and Taxonomy, 1st edition, Cichlid Press, 360 pages
Teugels G.G., and Thys van den Audenarde D.F.F., 1990, Description of a new species of Bryconaethiops (Teleosti; Characidae) from Nigeria and Cameron, Ichthyogical Exploration of Freshwaters, 1(3): 207 - 212
Thorpe R.S., 1987, Geographic variation: A synthesis of cause, data pattern and congruence in relation to subspecies, multivariate analysis and phylogenies, Bulletin of Zoology, 54: 3-11
http://dx.doi.org/10.1080/11250008709355549



. PDF(690KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Elagba H. A. Mohamed
. Wigdan A.S. Al-Awadi
Related articles
. Hydrocynus
. H. vittatus
. H. brevis and H. forskalii
. Length-weight Relationship
. Morphology
. Taxonomy
Tools
. Email to a friend
. Post a comment