The Nutrient and Carbon Dynamics that Mutually Benefit Coral and Seagrass in Mixed Habitats under the Influence of Groundwater at Bise Coral Reef, Okinawa, Japan 
Tomihiko Higuchi1,4  ,
Kimberly K. Takagi2,5 ,
Kana Matoba2 ,
Syusei Kobayashi1 ,
Ryota Tsurumi1 ,
Seiji Arakaki2,6 ,
Yoshikatsu Nakano3 ,
Hiroyuki Fujimura1
,
Kimberly K. Takagi2,5 ,
Kana Matoba2 ,
Syusei Kobayashi1 ,
Ryota Tsurumi1 ,
Seiji Arakaki2,6 ,
Yoshikatsu Nakano3 ,
Hiroyuki Fujimura1  ,
Tamotsu Oomori1 ,
Makoto Tsuchiya1
,
Tamotsu Oomori1 ,
Makoto Tsuchiya1
1. Department of Chemistry, Biology and Marine Science, University of the Ryukyus, 1 Senbaru, Nishihara, Okinawa 903-0213, Japan
2. Graduate School of Engineering and Science, University of the Ryukyus, 1 Senbaru, Nishihara, Okinawa 903-0213, Japan
3. Sesoko Station, Tropical Biosphere Research Center, University of the Ryukyus, 3422 Sesoko, Motobu, Okinawa 905-0227, Japan
4. Graduate School of Science and Technology, Shizuoka University, 836 Ohya, Surugaku, Shizuoka, 422-8529, Japan
5. Department of Marine Science, University of Georgia, 325 Sanford Drive, Athens, Georgia, 30602, USA
6. Amakusa Marine Biological Laboratory, Kyushu University, 2231 Tomioka, Reihoku, Amakusa, Kumamoto 863-2507, Japan
Author Correspondence author
International Journal of Marine Science, 2014, Vol. 4, No. 1 doi: 10.5376/ijms.2014.04.0001
Received: 27 Oct., 2013 Accepted: 29 Nov., 2013 Published: 04 Jan., 2014
2. Graduate School of Engineering and Science, University of the Ryukyus, 1 Senbaru, Nishihara, Okinawa 903-0213, Japan
3. Sesoko Station, Tropical Biosphere Research Center, University of the Ryukyus, 3422 Sesoko, Motobu, Okinawa 905-0227, Japan
4. Graduate School of Science and Technology, Shizuoka University, 836 Ohya, Surugaku, Shizuoka, 422-8529, Japan
5. Department of Marine Science, University of Georgia, 325 Sanford Drive, Athens, Georgia, 30602, USA
6. Amakusa Marine Biological Laboratory, Kyushu University, 2231 Tomioka, Reihoku, Amakusa, Kumamoto 863-2507, Japan
Author Correspondence author
International Journal of Marine Science, 2014, Vol. 4, No. 1 doi: 10.5376/ijms.2014.04.0001
Received: 27 Oct., 2013 Accepted: 29 Nov., 2013 Published: 04 Jan., 2014
© 2014 BioPublisher Publishing Platform
This is an open access article published under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Preferred citation for this article:
Higuchi et al, 2014, The Nutrient and Carbon Dynamics that Mutually Benefit Coral and Seagrass in Mixed Habitats under the Influence of Groundwater at Bise Coral Reef, Okinawa, Japan, International Journal of Marine Science, Vol.4, No.1 1-15 (doi: 10.5376/ijms.2014.04.0001)
Abstract
The coral species, Montipora digitata and seagrass, Thalassia hemprichii, co-inhabit the southern portion of the reef moat in Bise, Okinawa, Japan. To elucidate the biogeochemical relationship between coral and seagrass in mixed communities of the coral reef ecosystem, the carbon metabolisms and the inorganic nitrogen flux rates were estimated in various reef habitats. We used benthic chambers to investigate sandy, seagrass, coral-seagrass mixed communities, coral, and acorn worm habitats. Relatively high concentrations of nitrate and nitrite ions (NOx) were observed in all habitats due to coastal groundwater inflow. The uptake rate constant of NOx was the highest in the coral-seagrass habitat and was significantly different from the rate constant in the seagrass habitat, indicating that seagrass benefits from co-inhabitation with coral. Dissolution of CaCO3 was observed in the seagrass and coral-seagrass communities. This decline in basal coral carbonate substrate may contribute to increased fragmentation and dispersal of the coral habitat. On a biogeochemical scale, the coral-seagrass relationship benefits the seagrass in terms of NOx availability and benefits the coral in terms of carbonate dissolution, increasing fragmentation, and furthering habitat development.
Keywords
Carbon metabolism; Inorganic nitrogen flux; Coral-seagrass mixed community; Coral reef; Benthic chamber
1 Introduction
The biogeochemical dynamics between coral and seagrass ecosystems are important components of ecosystem management. Understanding these dynamics is especially important in coastal areas where increasing populations threaten the economic services these ecosystems provide. In order to understand broad-scale ecosystem function, recent management concerns have addressed the importance of functional group dynamics (Bellwood et al., 2004). Naeem (1998) stated that in order for ecosystems to resist “failure” after a disturbance event, species richness within functional groups is critical in maintaining ecosystem stability and reliability. For example, coral reefs serve as physical buffers for oceanic currents and waves, creating, over geologic time, a suitable environment for seagrass beds. In addition to these physical interactions, there are several biological and biogeochemical interactions between these interconnected ecosystems (Moberg and Folke, 1999). Seagrass beds interrupt freshwater discharge, are sinks for organic and inorganic materials as well as pollutants, and can generate an environment with the appropriate nutrient levels that promotes the growth of coral reefs (Miyajima et al., 2001; Umezawa et al., 2002).
If seagrass and coral in mixed habitats establish mutually symbiotic functional groups, it is possible that the presence of both can create an ecosystem that is more resilient to disturbance. However until recently, research has generally focused on the nutrient and carbon dynamics within these two ecosystems separately (Badgely et al. 2006, Grover et al., 2003; Marbà et al., 2006; Ohde and van Woesik, 1999; Tenore, 1988), and while few, if any, have addressed cases in which coral and seagrass co-inhabit the same area.
Ninomiya et al. (2006) suggested that there is a physically mutualistic relationship between coral and seagrass in mixed habitats. The study asserts that the vertical and horizontal entwining of seagrass stems and coral branches both on and under the surface of the seafloor gives stability to seagrass beds. Jompa and McCook (2003) also showed that in the case of canopy forming macrophytes, understory corals are often protected from bleaching damage by shading. This implies that a dense seagrass community formed around fragmented coral can encourage coral survival by providing protection from bleaching.
Manzellos et al. (2012) suggested that coral calcification rates are higher near seagrass beds, because seagrasses draw down CO2 in primary production and can store the carbon as biomass in their root systems. Thus, there is likely a biogeochemical relationship between seagrass and coral. Through the analysis of carbon and inorganic nitrogen dynamics, this study aims to elucidate this biogeochemical relationship. In Bise, Okinawa, Japan, there is a high population density of co-inhabiting seagrass and corals. Here, the southern portion of this reef moat is co-inhabited by the coral species, Montipora digitata and the seagrass species, Thalassia hemprichii. By comparing the biogeochemical interactions in the sand, seagrass, coral, and acorn worm habitats, with those in the coral-seagrass mixed habitat, we could identify the roles that each component has in the carbon and inorganic nitrogen dynamics both separately and interactively within the coral reef ecosystem.
2 Materials and Methods
2.1 Study site
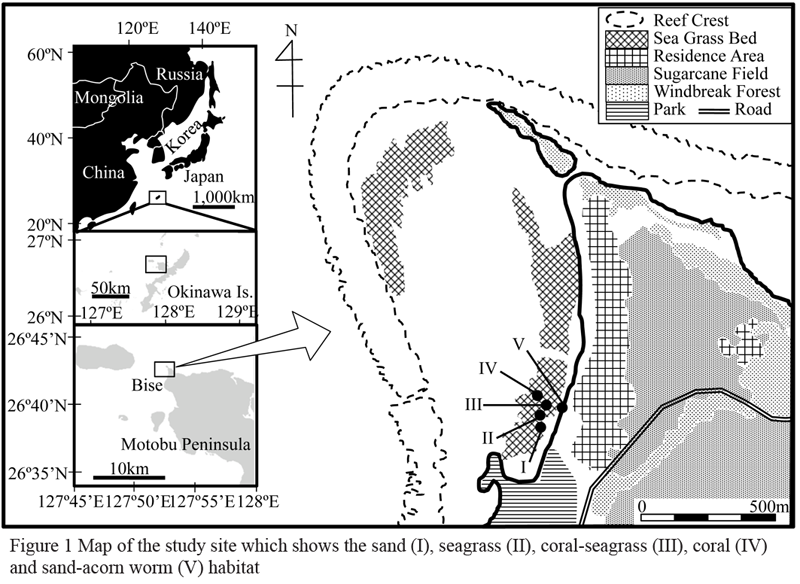
This study was conducted in the subtropical reef moat off of Bise Coast, Okinawa, Japan, from Aug. 1 to 6, 2008 (Figure 1). It is characterized by coral, seagrass, and coral-seagrass mixed communities. The dominant coral species is Montipora digitata, and the dominant seagrass species is Thalassia hemprichii. The most conspicuous benthic organisms inhabiting this area are the acorn worms; Schizocardium sp. and Ptychodera flava. These invertebrates are bioturbators which produce fecal casts on the sediment surface while burrowing into the coral reef sands (up to 24 indivduals m-2, pers. obsv.). The underlying substrate of these habitats is primarily dominated by limestone.
|
Figure 1 Map of the study site which shows the sand (I), seagrass (II), coral-seagrass (III), coral (IV) and sand-acorn worm (V) habitat |
2.2 Chamber experiment
Benthic chambers (Ishikawa et al., 2007) were used to conduct 2 h incubations in each habitat (Figure 2). Using gardening stands fitted with plastic bags as incubation chambers, the nutrient and carbon dynamics of five benthic habitats were assessed: sand only (SD, 100% sand), seagrass only (SG, 100% T. hemprichii), coral-seagrass (CS, T. hemprichii : 50%, M. digitata : 45%, sand : 5%), coral only (CR, M. digitata : 95%, sand : 5%), and acorn worm habitats (AC, 100% sand with 2-4 acorn worms). Three incubation chambers were placed in each habitat and water samples were collected from each chamber. For the SG, CS, and CR habitats, 44 L benthic chambers (1479 cm2 base area, 40 cm ht.) were deployed. Smaller chambers were used for the SD and AC habitats, 15 L benthic chambers (984 cm2 base areas, 20 cm ht.) to prevent the chamber from emerging out of the water during low tide. Before beginning the incubations, each habitat was cleared of benthic macroalgae, snails and other visible macrofauna to minimize other variables that could affect the inorganic nitrogen and carbon dynamics. In the SD habitat, the contributions of phytoplankton and/or other microorganisms in benthos and water column were estimated as a control.
|
Figure 2 Schematic diagram of the benthic chamber. First, hard flames are put on each habitat (1). Then, clear plastic bags were covered on the flames (2). And then, hard flames was put again on the plastic bags (3). Water samples are taken by using syringes before and after the 2-h chamber incubations (4) |
.png)
2.3 Chemical and environmental parameters
Seawater samples were collected by syringe at the beginning and end of each of the incubations. The pH and total alkalinity (TA) were determined using a pH electrode (pH meter Orion 4-star, Thermo-Fisher Scientific) on the NBS scale and the Gran plot method using a total alkalinity titrator (ATT-05, Kimoto), respectively. Accuracy of these measurement were ±0.05 for pH and ±10 µmol kg-1 for TA. Salinity (S) was measured by a salinometer (Portsal 8410A, Guildline) based on comparison with IAPSO standard water (S = 34.993). Ammonium, nitrate, and nitrite ions were measured by an automatic water analyzer (QuAAtro, BRAN+LUEBBE).
Data loggers (Water Temp Pro v2 Data Logger, HOBO) and photon sensors (MDS-MkV/L, Alec Electronics Co.) were deployed beside the chamber to record temperature and photon flux. Tidal level data were provided by the Okinawa Meteorological Observatory.
2.4 Groundwater inflow
Groundwater comes out of sediment through the limestone. To examine the effects of groundwater outflow into the reef moat, the initial total alkalinity (TA), nitrate + nitrite (NOx), and ammonium (NH4+) concentrations during the lowest spring tides (1-Aug, 12:00, 1-Aug, 16:00, and 4-Aug, 16:00) of the sampling period were plotted against salinity (S). The slope of each regression line (Rslope) was used to calculate the change in concentration (∆Cinflow) due to groundwater inflow during the chamber incubation.
∆Cinflow = Rslope • ∆Sfinal-initial (Eq.1)
where ∆Cinflow is the inflow concentration of total alkalinity, NOx or NH4+, Rslope is the slope of each regression line in Figure 3, ∆Sfinal-initial is the difference in salinity between final and initial samples in each incubation chamber.
2.5 Data analysis
The difference between chemical constituents in each chamber over a given period was attributed to biological activity. However, as we detected a decrease in salinity during the incubations, most of the chambers in this study were influenced by groundwater inflow. Therefore, we accounted for the effect of inflowing water to estimate the biological activity as follows:
∆Cbiological = Cfinal - Cinitial - ∆Cinflow (Eq.2)
Where, ∆Cbiological is the change in TA, DIC, NOx, or NH4+ during an incubation due to the biological activity within a given chamber.
Using the ∆Cbiological of total alkalinity (TA) and total dissolved inorganic carbon (DIC) data, the rates of organic carbon production (OP, mmol m–2 h–1) and inorganic carbon production (IP, mmol m–2 h–1) were calculated using the alkalinity anomaly technique (Smith 1973, Smith and Kinsey 1978). IP was expressed by using: the changes in TA, volume of the chamber (V), base area of the chamber (A) and incubation time (t), as follows:
Using the ∆Cbiological of total alkalinity (TA) and total dissolved inorganic carbon (DIC) data, the rates of organic carbon production (OP, mmol m–2 h–1) and inorganic carbon production (IP, mmol m–2 h–1) were calculated using the alkalinity anomaly technique (Smith 1973, Smith and Kinsey 1978). IP was expressed by using: the changes in TA, volume of the chamber (V), base area of the chamber (A) and incubation time (t), as follows:
IP=-ΔTAbiological⋅V/2⋅A⋅t (Eq. 3)
OP was calculated by using the changes in total dissolved inorganic carbon (DIC), volume of chamber (V), the base area of chamber (A), incubation time (t) and IP, as follows:
OP=(-ΔDICbiological⋅V/A⋅t)−IP (Eq. 4)
International Journal of Marine Science
• Volume 4



View Options
. PDF(830KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Tomihiko Higuchi
. Kimberly K. Takagi
. Kana Matoba
. Syusei Kobayashi
. Ryota Tsurumi
. Seiji Arakaki
. Yoshikatsu Nakano
. Hiroyuki Fujimura
. Tamotsu Oomori
. Makoto Tsuchiya
Related articles
. Carbon metabolism
. Inorganic nitrogen flux
. Coral-seagrass mixed community
. Coral reef
. Benthic chamber
Tools
. Email to a friend
. Post a comment
. PDF(830KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Tomihiko Higuchi
. Kimberly K. Takagi
. Kana Matoba
. Syusei Kobayashi
. Ryota Tsurumi
. Seiji Arakaki
. Yoshikatsu Nakano
. Hiroyuki Fujimura
. Tamotsu Oomori
. Makoto Tsuchiya
Related articles
. Carbon metabolism
. Inorganic nitrogen flux
. Coral-seagrass mixed community
. Coral reef
. Benthic chamber
Tools
. Email to a friend
. Post a comment