The Effects of Temperature on Gene Expression in the Indo-Pacific Reef-building Coral Seriatopora hystrix: Insight from Aquarium Studies in Southern Taiwan 
Anderson B. Mayfield
1,2 
,
Yi-Hsuan Chen
3 
,
Chang-Feng Dai
3 ,
Chii-Shiarng Chen
1,4,5
1 National Museum of Marine Biology and Aquarium, 2 Houwan Rd., Checheng, Pingtung 944, Taiwan, R.O.C.
2 Living Oceans Foundation, 8181 Professional Place, Suite 215, Landover, MD 20785, United States of America
3 Institute of Oceanography, National Taiwan University, 10617 No.1, Sec. 4 Roosevelt Rd., Taipei 106, Taiwan, R.O.C.
4 Graduate Institute of Marine Biotechnology, National Dong-Hwa University, 2 Houwan Rd., Checheng, Pingtung 944, Taiwan, R.O.C.
5 Department of Marine Biotechnology and Resources, National Sun Yat-Sen University, 70 Lianhai Rd., Kaohsiung 804, Taiwan, R.O.C.
Author
Correspondence author
International Journal of Marine Science, 2014, Vol. 4, No. 50 doi: 10.5376/ijms.2014.04.0050
Received: 05 Jun., 2014 Accepted: 13 Jul., 2014 Published: 22 Aug., 2014
© 2014 BioPublisher Publishing Platform
This is an open access article published under the terms of the
Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Preferred citation for this article:
Mayfield et al., 2014, The effects of temperature on gene expression in the Indo-Pacific reef-building coral Seriatopora hystrix: insight from aquarium studies in Southern Taiwan, International Journal of Marine Science, Vol.4, No.50 1-22 (doi: 10.5376/ijms.2014.04.0050)
Abstract
Corals from upwelling reefs of Southern Taiwan have been shown to acclimate to both highly variable and abnormally elevated temperatures. To better understand the genetic basis for Taiwanese reef coral acclimation to such temperature regimes, eight and six genes hypothesized to be thermo-sensitive were targeted in specimens of the common stony coral Seriatopora hystrix and their endosymbiotic dinoflagellate (genus Symbiodinium) communities, respectively, from two reefs in Southern Taiwan exposed to two different temperature profiles. Coral specimens from Houbihu, a reef characterized by extensive, spring-tide upwelling, displayed few provocative gene expression changes when exposed for two days to a temperature they rarely encounter in situ, 30℃. In a second experiment, corals from Houbihu and Houwan, the latter being a reef characterized by more stable temperatures, were exposed to either a stable (26℃) or a fluctuating (23-29℃ over a 5-h period) temperature regime for seven days, and it was found that 50% of the genes demonstrated significant changes in expression across treatments, regardless of the site of origin. This suggests that exposure to a variable temperature for seven days may have a more dramatic effect on the sub-cellular behavior of this species than a two-day incubation at a stable, though potentially stress-inducing, temperature.
Keywords
Acclimation; Acclimatization; Coral reef, Dinoflagellate; Endosymbiosis; Gene expression; Temperature
Given the threats of global climate change (GCC) towards scleractinian corals and the reef ecosystems they construct, there is an urgent need to better understand their physiological (Gates and Edmunds, 1999) and sub-cellular (van Oppen and Gates, 2006) mechanisms of acclimatization to altered temperature regimes (Mayfield et al., 2013a). An emerging model for the study of GCC effects on reef corals is Seriatopora hystrix (Figure 1), which is amongst the most ubiquitously distributed scleractinians in the Indo-Pacific (Loh et al., 2001; Veron, 2000). S. hystrix has served as a model coral for research in diverse locations across the Pacific Rim, such as Australia (e.g., Bongaerts et al., 2011) and Taiwan (e.g., Fan et al., 2006; Putnam et al., 2010; Tung et al., 2006), and isamongst the most sensitive corals to environmental change (Loya et al., 2001). However, in Southern Taiwan, S. hystrix lives in relatively warm waters (summer mean = ~28.5℃) whose temperatures can change rapidly due not only to the tidal cycle (Meng et al., 2008), but also to episodic upwelling (Chen et al., 2004; Jan and Chen, 2008).
|
.png)
Figure 1 Map of Southern Taiwan’s Hengchun Peninsula, including the locations of the two study sites (Houwan and Houbihu), alongside an image of the target organism Seriatopora hystrix, which is common at both sites. For a detailed treatise on the temperature environment of the two sites, readers are pointed towards Mayfield et al. (2012a). Briefly, the average monthly temperature ranges of Houbihu and Houwan from 2009 to 2010 were 6.33±2.03 and 3.19±0.61?, respectively, and this represents a significant difference (student’s t-test, t=5.13, p<0.01). The size of the S. hystrix colony in the inset is approximately 15 cm in diameter
|
|
.png)
Table 1 Summary of the two Seriatopora hystrix experiments conducted at Taiwan’s National Museum of Marine Biology and Aquarium (NMMBA). Although the experiments were conducted a year apart, the same aquarium system was used to conduct each one. Also, the acclimation temperatures, which aimed to approximate in situ values at the time of coral collection, were slightly different for the two experiments due to in situ differences at the time of coral collection
|
S. hystrix is known to have limited dispersal capacity (Ayre and Hughes, 2000), a trait that may be associated with its ability to readily adapt to a diverse array of environments (Nunes et al., 2009; Sanford and Kelly, 2011). Indeed, prior work on S. hystrix populations from the thermally dynamic upwelling sites mentioned above found that neither the coral hosts nor their resident Symbiodinium populations mount a gene-level molecular chaperone (heat shock protein-70 [hsp70]) response when exposed to 30℃ for 48 h (Mayfield et al., 2011; i.e., the elevated temperature experiment [ETE]). This was hypothesized to be a stress-inducing temperature given that it is ~1℃ greater than the average summer temperature of Houbihu (Figure 1), the upwelling site (UWS) within Nanwan Bay (Taiwan’s southernmost embayment) from which the experimental corals were collected; briefly, previous studies (e.g., Coles and Brown, 2003; Hoegh-Guldberg and Smith, 1989) have found that extended exposure to temperatures >1℃ above the mean summer high can result in bleaching in many reef coral species.
Given these results, Mayfield et al. (2011) hypothesized that S. hystrix residing in the UWS may have special adaptations for life in thermodynamically variable environments, as the temperature at the UWS can change by up to 9℃ in a single day during spring-tide upwelling events that occur throughout the year (but mainly in the northern summer; Mayfield et al., 2012a). Mayfield et al. (2012a, 2013c) attempted to test this hypothesis with a laboratory-based reciprocal transplant in which corals from this UWS were exposed to either a variable (23-29℃ over a 5-h period) or stable (26℃) temperature treatment while conspecifics from a non-upwelling site (i.e., NUWS) characterized by relatively stable (over diel and annual timescales) temperatures, Houwan (Figure 1), were exposed to the same two temperature regimes for seven days. It was hypothesized that corals transplanted to a “foreign” temperature regime would be physiologically compromised in this variable temperature experiment (VTE), though this hypothesis was confirmed only upon assessment of coral growth (Mayfield et al., 2012a); other parameters appear to have been more significantly influenced by the temperature regime alone.
To gain more insight into the genetic basis underlying the phenotypic plasticity of these Taiwanese S. hystrix populations, expression of 14 gene mRNAs whose respective proteins and cellular pathways have been hypothesized to be disrupted upon exposure to elevated temperature was measured in samples of the ETE and VTE (Table 1); these included mRNAs encoding proteins involved in photosynthesis, metabolism, osmoregulation, the cytoskeleton, and the stress response (see the Materials and Methods for detailed rationale for choosing the respective genes.). Gene expression was hypothesized to remain similar over time and between treatments in samples from the UWS exposed to 30℃for two days in the ETE given the lack of hsp70 modulation documented at these temperatures by Mayfield et al. (2011). In contrast, it was hypothesized that the majority of the genes would be expressed at different levels between the stable and variable temperature treatments in the VTE in corals from the UWS only, given that exposure to a familiar temperature change can drive significant changes in mRNA expression in other marine organisms (e.g., Gracey et al., 2008). It was further hypothesized that corals from the UWS would demonstrate higher levels of target gene expression than conspecifics from the NUWS. Briefly, corals inhabiting variable temperature environments could be expected to express higher mRNA levels; the reasoning behind this is due to the need for rapid increases in protein translation at times at which temperature increases have incapacitated the standing pool of intracellular proteins in response to elevated temperature-induced protein denaturation (Hazel and Prosser, 1974). This is a strategy employed by not only intertidal limpets (Dong et al., 2008), but also other corals (Barshis et al., 2013).
1 Results
1.1 ETE: Symbiodinium gene expression
Recovery of the Solaris™ spike (Thermo-Scientific) was similar between treatments and over time (data not shown) in samples of the ETE (Table A1), all 24 of which were found to be composed only of Symbiodinium of clade C (Ct<30; Ct>35 for clade A and D assays). Symbiodinium gene expression was normalized to both spike recovery and the Symbiodinium genome copy proportion (GCP; Figure 2A); the latter of which was also found to be similar across treatments and time (Table A1). Expression of ribulose-1,5-bisphosphate carboxylase-oxygenase large subunit (rbcL; Figure 2B), photosystem I (subunit III; psI; Figure 2C), phosphoglycolate phosphatase (pgpase; Figure 2D), nitrate transporter-2 (nrt2; Figure 2E), and ascorbate peroxidase (apx1; Figure 2F) was similar over time and between treatments (Table A1).
|

Table A1 Symbiodinium exogenous RNA spike recovery, genome copy proportion (GCP), and gene expression results from the Seriatopora hystrix elevated temperature experiment (ETE). Repeated-measures ANOVAs were conducted with JMP using a MANOVA model as described in the text. The results from the S. hystrix variable temperature experiment (VTE) are presented in the right-most column. “UWS” = upwelling site (Houbihu). “NUWS” = non-upwelling site (Houwan)
|
|
.png)
Figure 2 ETE: Symbiodinium gene expression. Symbiodinium genome copy proportion (GCP [unit-less]; A) and expression of five genes (B-F) in Seriatopora hystrix samples exposed to either a control (27?; hollow squares) or elevated (30?; filled diamonds) temperature for 48 h. Please see the text or Tables A1-3 for full gene names. Gene expression data were normalized as described in the text and presented as unit-less. In all panels, error bars represent standard error of the mean (n=3 replicates per treatment-time)
|
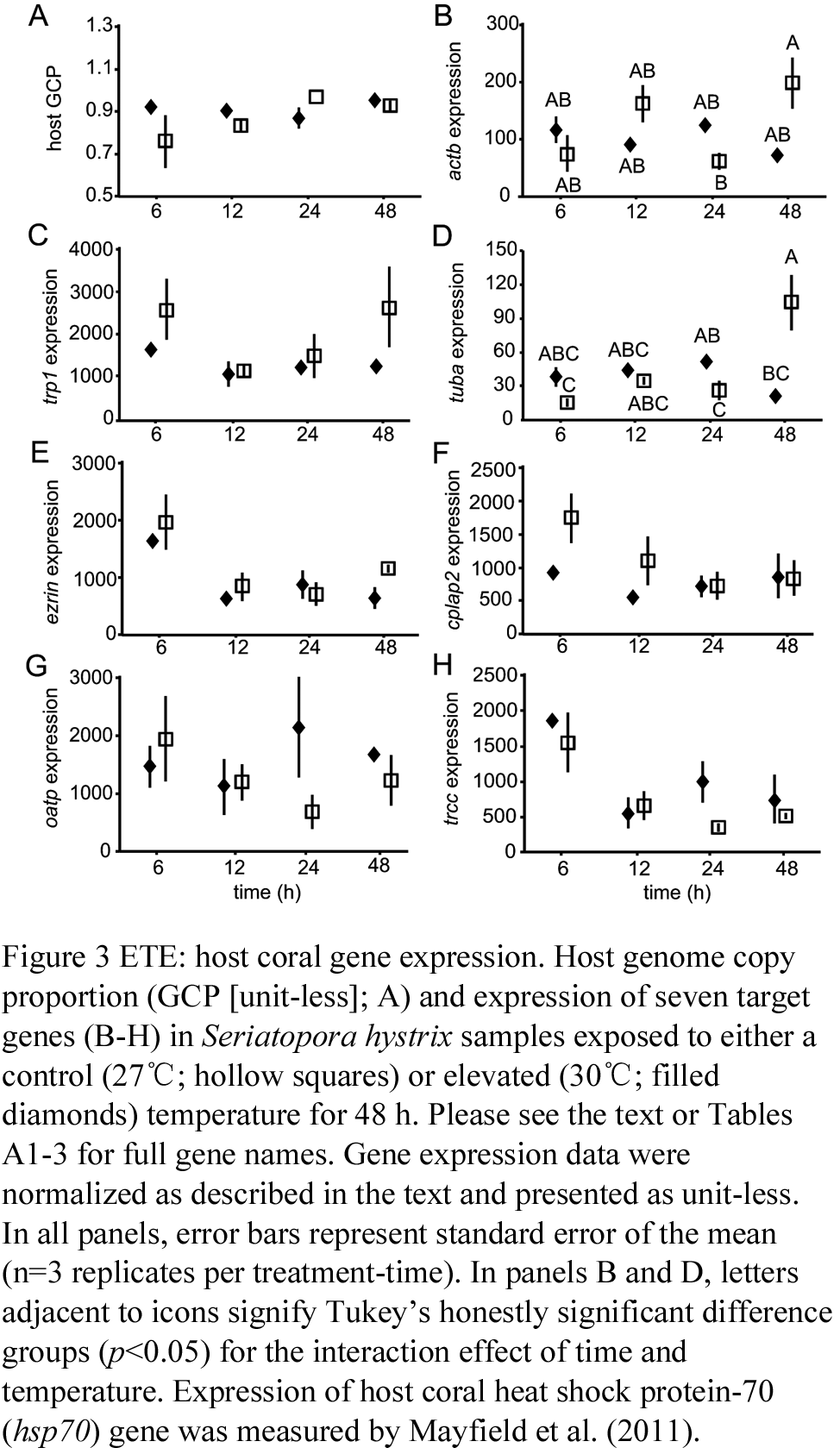
1.2 ETE: host coral gene expression
Expression of the eight host genes was normalized to recovery of the Solaris spike (data not shown) and the host GCP (Figure 3A); the latter of which was found to be stable over time and between treatments (Table A2). Only β-actin (actb; Figure 3B) and α-tubulin (tuba; Figure 3D) were significantly affected by the interaction of time and temperature; expression of the former in the control nubbins increased ~4-fold from the 24 to the 48 h sampling time (Figure 3B). In contrast, tuba was expressed at 2-fold higher levels in Figure 3 ETE: host coral gene expression. Host genome copy proportion (GCP [unit-less]; A) and expression of seven target genes (B-H) in Seriatopora hystrix samples exposed to either a control (27℃; hollow squares) or elevated (30℃; filled diamonds) temperature for 48 h. Please see the text or Tables A1-3 for full gene names. Gene expression data were normalized as described in the text and presented as unit-less. In all panels, error bars represent standard error of the mean (n=3 replicates per treatment-time). In panels B and D, letters adjacent to icons signify Tukey’s honestly significant difference groups (p<0.05) for the interaction effect of time and temperature. Expression of host coral heat shock protein-70 (hsp70) gene was measured by Mayfield et al. (2011). the high temperature samples of time 24 relative to the controls. Then, at sampling time 48, expression of this gene was 5-fold higher in controls. Tropomyosin (
trp1;
Figure 3C),
ezrin (
Figure 3E), phospholipase-
α2 (
cplap2;
Figure 3F), organic anion transporter (
oatp;
Figure 3G), and transient receptor cation channel (
trcc;
Figure 3H) were, on the other hand, expressed at similar levels between treatments across the four sampling times (
Table A2).
|
.png)
Figure 3 ETE: host coral gene expression. Host genome copy proportion (GCP [unit-less]; A) and expression of seven target genes (B-H) in Seriatopora hystrix samples exposed to either a control (27?; hollow squares) or elevated (30?; filled diamonds) temperature for 48 h. Please see the text or Tables A1-3 for full gene names. Gene expression data were normalized as described in the text and presented as unit-less. In all panels, error bars represent standard error of the mean (n=3 replicates per treatment-time). In panels B and D, letters adjacent to icons signify Tukey’s honestly significant difference groups (p<0.05) for the interaction effect of time and temperature. Expression of host coral heat shock protein-70 (hsp70) gene was measured by Mayfield et al. (2011)
|
|

Table A2 Host genome copy proportion (GCP) and gene expression results from the Seriatopora hystrix elevated temperature experiment (ETE). Repeated-measures ANOVAs were conducted with JMP using a MANOVA model as described in the text. Statistically significant differences are underlined. The results from the S. hystrix variable temperature experiment (VTE) are presented in the right-most column
|
1.3 VTE: Symbiodinium gene expression
1.4 VTE: host coral gene expression
The expression of both actb (Figure 5C) and cplap2 (Figure 5G) was affected by the experimental treatments (Table A3); the former was found to be 30% higher in samples exposed to the variable temperature regime. cplap2 was characterized by an interaction effect in which corals from the NUWS exposed to the variable temperature regime expressed 2.1-fold higher levels than corals from the UWS exposed to this same temperature profile. Of the 14 target genes assessed across both compartments, 2 (host actb and tuba; 14%) were significantly affected by temperature in the ETE, whereas 7 (Symbiodinium rbcL, psI, pgpase [from Mayfield et al., 2012a], and hsp70 and host coral hsp70, actb, and cplap2; 50%) demonstrated significant differences between temperature regimes in the VTE. This difference in the overall proportion of target genes demonstrating a significant difference across the two experiments was marginally significant (X2=3.9, p=0.05).
1.5 VTE: host coral genotyping
With the exception of Sh4-001, all microsatellite loci were polymorphic (Table 2); the former was excluded from the calculations. The number of alleles (NA), observed heterozygosity (Ho), and expected heterozygosity (He) for each remaining locus ranged from 1 to 7 (mean=3.9), 0 to 0.83 (mean=0.53), and 0 to 0.89 (mean=0.63), respectively. Deviations from Hardy-Weinberg equilibrium and heterozygote deficiencies were suggested by inbreeding coefficient (FIS) values, which were significant for samples from the UWS (FIS=0.24, p<0.05), but not the NUWS (FIS=0.099, p=0.20). The overall FIS value of 0.16 was statistically significant (p<0.05), suggesting that inbreeding mayexist within these two populations. There were no null alleles across the seven loci (data not shown). Finally, no significant genetic differentiation was found between the Houbihu and Houwan samples (fixation index [FST]=-0.012, p>0.05), though this could be partially attributed to the small sample size (n=6 for each site of origin).
|
.png)
Table 2 Host coral genotyping results. “N” = sample size. “NA” = number of alleles. “NPA” = number of private alleles. “Ho” = observed heterozygosity. “He” = expected heterozygosity. “FIS” = inbreeding coefficient. “UWS” = upwelling site (Houbihu). “NUWS” = non-upwelling site (Houwan)
|
Expression of nrt2 (Figure 4A) and heat shock protein-70 (hsp70; Figure 4B) was assessed in Symbiodinium populations within the 24 samples, all of which were shown to be of clade C only by real-time PCR (Ct<30; Ct>35 for clade A and D assays). Although there were no significant effects of temperature, site of origin, or their interaction on expression of the former (Table A3), expression of Symbiodinium hsp70 was significantly higher (1.8-fold) in samples exposed to a variable temperature regime (Table A3). The influence of the temperature regime had a significantly greater influence on Symbiodinium gene expression in samples of the VTE (4/6 of the target genes were significantly affected by temperature when combining the results herein with the three genes [psI, pgpase, and rbcL] assessed by Mayfield et al. [2012a].) versus those of the ETE (0/6; X2=7.6, p<0.01).
|
.png)
Figure 4 VTE: Symbiodinium gene expression. nrt2 (A) and hsp70 (B) gene expression from Symbiodinium within Seriatopora hystrix nubbins from Houbihu (“upwelling” site [UWS]) or Houwan (“non-upwelling” site [NUWS]) exposed to either a stable (hollow squares) or variable (filled diamonds) temperature treatment for seven days. Please see the text or Tables A1-3 for full gene names. Gene expression data were normalized as described in the text and presented as unit-less. Error bars represent standard error of the mean in all panels (n=3 replicates [6 pseudo-replicates] per treatment-site of origin), and in panel B only, the letters adjacent to icons denote Tukey’s honestly significant difference groups (p<0.05) for the temperature effect only
|
|

Table A3 Symbiodinium and host coral gene expression results from the Seriatopora hystrix variable temperature experiment (VTE). Two-way ANOVAs were conducted with JMP as described in the text. Statistically significant differences are underlined. “HSD” = honestly significant difference
|
Expression of the stress-targeted gene hsp70 (Figure 5A), four cytoskeleton-targeted genes; actb (Figure 5C), trp1 (Figure 5D), tuba (Figure 5E), and ezrin (Figure 5F), and three osmoregulation-targeted genes; cplap2 (Figure 5G), oatp (Figure 5H), and trcc (Figure 5I), were normalized to both recovery of the Solaris spike (data not shown) and the host GCP; the latter of which was shown to be stable between sites of origin and temperature treatments (data not shown).
Although host hsp70 expression was found at significantly higher (~3.5-fold) levels in samples exposed to the stable temperature regime (Figure 5A), there was neither site of origin nor interaction effects (Table A3). There was also no significant correlation between Symbiodinium and host hsp70 expression (Figure 5B; linear regression t-test, t=2.1, p>0.05).
|
.png)
Figure 5 VTE: host coral gene expression. Expression of eight host coral genes; hsp70 (A), actb (C), trp1 (D), tuba (E), ezrin (F), cplap2 (G), oatp (H), and trcc (I), and correlations in hsp70 expression within the holobiont (B) in Seriatopora hystrix nubbins from Houbihu (“upwelling” site [UWS]) or Houwan (“non-upwelling” site [NUWS]) exposed to either a stable (hollow squares of panels A and C-I) or variable (filled diamonds of panels A and C-I) temperature for seven days. Please see the text or Tables A1-3 for full gene names. Gene expression data were normalized as described in the text and presented as unit-less. Error bars represent standard error of the mean (n=3 replicates [6 pseudo-replicates] per treatment-site of origin). In panels A and C, letters adjacent to icons correspond to Tukey’s honestly significant difference (HSD) temperature treatment groups only (p<0.05), while letters adjacent to icons in panel G correspond to the interaction effect of site of origin and temperature treatment (Tukey’s HSD, p<0.05)
|
1.6 Site of origin comparisons
Upon pooling data across experiments (Figure 6), the Symbiodinium GCP (data not shown) was found to be similar between sites of origin (student’s t-test, t=0.91, p=0.37). On the other hand, rbcL expression was significantly higher (16-fold) in samples from the UWS relative to those of the NUWS (Wilcoxon Z=3.5, p<0.001). Although expression of psI was statistically similar between the two sites (Z=1.3, p=0.19), pgpase was expressed at 2.0-fold higher levels in corals of the UWS (Z=4.7, p<0.001). In contrast, nrt2 was expressed at similar levels across sites (student’s t=0.74, p=0.46), as was apx1 (Wilcoxon Z=1.8, p=0.076). Finally, Symbiodinium hsp70 was expressed at signi- ficantly higher (4.1-fold) levels in corals from the NUWS relative to those of the UWS (Z=3.6, p<0.001).
|
.png)
Figure 6 Global site of origin differences in gene expression. To compare the average expression of each of the 14 target genes across the two sites of origin, Houbihu (upwelling site [UWS]; speckled columns) and Houwan (non-upwelling site [NUWS]; hatched columns), data from the 12 UWS samples of the Seriatopora hystrix variable temperature experiment (VTE) and all 24 samples of the elevated temperature experiment (ETE) were compared against the expression data from the 12 NUWS samples from the VTE. Please see the text or Tables A1-3 for full gene names. Gene expression values were first converted to fold changes relative to the lowest expression sample within the 48 assessed. Then, mean fold changes were log-transformed in order to present all data on the same scale. Asterisks (“*”) denote significant differences detected by Bonferroni-adjusted Wilcoxon rank-sum tests, and error bars represent standard error of the mean (n=36 and 12 for the UWS and NUWS, respectively). The statistically significant difference in host coral oatp expression between sites must be interpreted with caution, as different primer concentrations were used between experiments (Table 4). “hsp70z” and “hsp70h” refer to the hsp70 homolog from the Symbiodinium and host coral compartments, respectively
|
With respect to the host coral genes, actb was expressed at similar levels between sites (Wilcoxon Z=0.20, p=0.84). On the other hand, trp1 was expressed at 41-fold higher levels in corals of the UWS (Z=3.0, p<0.01), and tuba was expressed at 2.3-fold higher levels in those from the NUWS (Z=3.5, p<0.001). ezrin and cplap2 were expressed at significantly higher (9.4 and 320-fold, respectively) levels in samples from the UWS relative to those from the NUWS (Z=2.6 and 2.8, p=0.01 and <0.01, respectively), as was oatp (140-fold difference; Z=2.3, p<0.05, though see caveats in caption for Figure 6).
The osmoregulation gene trcc was expressed at 3.1-fold higher levels in corals from the UWS (Z=2.3, p<0.05). Finally, the host molecular chaperone hsp70 was expressed at significantly lower (1.8-fold) levels in corals from the UWS relative to those of the NUWS (Wilcoxon Z=2.5, p<0.05). Although 7/14 (50%) of the genes were more highly expressed in samples from the UWS versus only 3/14 (21%) of the genes demonstrating higher expression in samples from the NUWS, this difference in proportions was not statistically significant (X2=2.5, p=0.11).
2 Discussion
From a collective assessment of the data obtained herein and in prior works (Table 3), it appears that S. hystrix colonies from Houbihu (UWS), Southern Taiwan do not demonstrate a pronounced mRNA-level response to a temperature hypothesized to induce stress, 30℃.Specifically, expression of none of the six Symbiodinium genes spanning three cellular processes; photosynthesis, metabolism, and the stress response, differed between treatments at any sampling time in the ETE. On the other hand, expression of host tuba decreased 2-fold from the 24 to the 48 h sampling time in the high temperature samples. Although such a decrease could provide evidence in support of the hypothesis that the coral cytoskeleton may become compromised under potentially stress-inducing conditions (Mayfield and Gates, 2007), a similar change in only one other target gene was observed; actb was also expressed at higher levels in controls relative to high temperature samples at the final sampling time, though this difference was not statistically significant. It should be noted that actb was the only target gene that was affected by temperature in both experiments, discounting its utility as a housekeeping gene for this coral species. However, the difference in expression between samples of the two temperature regimes was only ~30% in the VTE.
|
.png)
Table 3 Summary of results. No coral colonies of the elevated temperature experiment (ETE) or nubbins of the variable temperature experiment (VTE) died over the course of the experiments, and for the ETE, in which only a small proportion of the 18 colonies was sacrificed for molecular analyses, the remaining colonies survived under ambient conditions at NMMBA for at least six months after the termination of the experiment
|
This absence of a temperature-driven response in 12/14 target genes across both compartments of the holobiont in samples exposed to 30℃ may suggest that exposure to a stable, elevated temperature regime does not elicit a stress response in specimens of S. hystrix from Southern Taiwan. Exposure to 30℃ has typically been shown to result in stress in S. hystrix elsewhere (Hoegh-Guldberg and Smith, 1989; Loya et al., 2001), and this should manifest at the gene level over the course of only several hours (Feder, 1996). Although it is tempting to speculate that this lack of an mRNA-level response to exposure to 30℃ in none of the six Symbiodinium target genes, and all but two (actb and tuba) of the eight host coral gene targets, stems from adaptations to life in a fluctuating temperature environment (Mayfield et al., 2011), a targeting of a greater number of gene and protein candidates over a longer sampling time will be necessary to demonstrate that these corals indeed display no signs of a sub-cellular stress response upon exposure to this temperature; indeed, <0.1% of the transcriptome of the S. hystrix-Symbiodinium holobiont was queried herein, and current efforts employing next generation sequencing-based technology (Mayfield et al., in prep.) seek to assess the whole-transcriptome response of these samples to gain further insight into the molecular mechanisms by which S. hystrix acclimates to altered temperature regimes in the laboratory and acclimatizes to them in the upwelling reef ecosystems in which they reside.
Additionally, in order to determine whether the absence of a gene-level is related to thermal history, a static ETE would also need to be conducted with conspecifics from the NUWS; such an experiment would help to strengthen the hypothesis put forth in prior works (e.g., Oliver and Palumbi, 2011; Mayfield et al., 2013b) that corals from variable temperature environments perform better at elevated temperatures than those from stable temperature habitats. If such a differential response were indeed documented, it would be attributable to acclimatization rather than adaptation, as the UWS and NUWS populations were found to be genetically identical (Table 2) and possess similarly homogenous Symbiodinium assemblages. That said, neutral markers, such as the microsatellites employed herein, cannot detect genetic differences driven by environmental changes (Foret et al., 2007) and so future studies seeking to uncover genetic differences underlying phenotypic plasticity should not only utilize larger sample sizes but also analyze genes that could be hypothesized to be under selection.
In contrast to incubation at a static, elevated temperature for two days, exposure to a variable temperature regime for seven days had a dramatic influence on gene expression; 50% of the genes were significantly affected by temperature in the VTE. This was mainly driven by the Symbiodinium gene expression profiles; although no Symbiodinium genes were significantly influenced by a 2-day exposure to 30℃, four of the six targets (67%); rbcL, psI, pgpase (assessed in Mayfield et al., 2012a), and hsp70 (assessed herein), were significantly influenced by temperature regime in the VTE. This was a significantly higher proportion, suggesting that exposure to a variable temperature regime over seven days had a more dramatic influence on the Symbiodinium mRNA-level response than exposure to a static, elevated one for two days. Mayfield et al. (2012a) took the elevated FV/FM values in these same samples to indicate that corals exposed to variable temperature may have a high capacity for photosynthesis, though an analysis of the degree of carbon fixation (sensu Furla et al., 2000) will be necessary to demonstrate whether such mRNA concentration and FV/FM increases actually lead to higher photosynthetic output of the resident clade C Symbiodinium populations.
The reason why exposure to a familiar, variable temperature regime had such a strong effect on Symbiodinium gene expression may be due to the need to rapidly adjust levels of protein translation in unstable environments. It has recently been found that corals may “front-load” gene mRNAs (Mayfield et al., 2011; Barshis et al., 2013), whereby expression is constitutively elevated at relatively high levels due to potentially high protein translation demands stemming from life in unstable environments. This strategy is commonly employed by intertidal invertebrates (Somero, 2010), such as limpets (Dong et al., 2008), and could explain the high levels of photosynthesis gene expression in the Symbiodinium populations within samples exposed to a variable temperature regime. mRNA front-loading theoretically allows for faster rates of protein translation that may be necessary when the standing pool of proteins becomes denatured due to abrupt temperature changes, such as the dramatic thermal shifts associated with upwelling events (Hochachka and Somero, 2002). By maintaining high levels of mRNA expression during such periods, corals and their resident Symbiodinium populations would have a higher scope for translation of proteins that may have been compromised by dramatic increases or decreases in temperature. This front-loading phenomenon may also explain why 7 of the 14 target genes were expressed at significantly higher levels in corals from the UWS, Houbihu, while only 3 were expressed at higher levels in those of the NUWS, Houwan (Figure6), a site characterized by a considerably more stable temperature profile (Mayfield et al., 2012a).
On the other hand, two of the three stress genes targeted herein and in a prior work (Mayfield et al., 2011), Symbiodinium and host coral hsp70, were instead expressed at higher levels in corals from the NUWS, suggesting that corals of the UWS do not necessarily front-load stress gene expression to a greater extent than conspecifics from the NUWS. As such, the front-loading mRNA loading phenomenon may be utilized for certain cellular response pathways, such as photosynthesis, but not all. A more thorough understanding of the behavior and function of transcription factors involved in the thermal acclimation response, such as heat shock factors (HSFs, Akerfelt et al., 2010), will ultimately aid in the development of an understanding of why the mRNA-level response differed between corals exposed to familiar or unfamiliar temperature regimes, as well as between sites. Such an analysis may also help to elucidate whether thermal history (e.g., prior exposure to upwelling) or the temperature regime itself is more important in driving the sub-cellular response of S. hystrix to temperature changes.
It is currently unclear whether such differences in gene expression between sites and temperature regimes documented herein and in previous works (e.g., Barshis et al., 2013) actually lead to an enhanced capacity for protein translation, as expression of the respective proteins was not measured in previous coral-based studies, nor was their activity. As such, it is hoped that both gene and protein expression, as well as activity of the latter, will be simultaneously measured in coral samples of future temperature manipulation studies such that the understanding of the molecular means by which corals respond to differential temperature regim
.png)
.png)
.png)
.png)
.png)


