Diet and Trophic Status of Fish Landed by Tropical Artisanal Bait Fishers, Mida Creek Kenya 
2. Currently at Rift Valley Water and Service Board PO Box 561-20117, Naivasha
3. Department of Biological Sciences, University of Nairobi PO Box 30197-00100, Nairobi, Kenya 4. Kenya Marine and Fisheries Research Institute, PO Box 81651, Mombasa, Kenya
 Author
Author  Correspondence author
Correspondence author
International Journal of Marine Science, 2015, Vol. 5, No. 42 doi: 10.5376/ijms.2015.05.0042
Received: 27 Mar., 2015 Accepted: 26 May, 2015 Published: 03 Jul., 2015
Kihia C .M. , Hendrick Y., Muthumbi A., Okondo J., Nthiga A and Njuguna V. M., 2015, Diet and trophic status of fish landed by tropical artisanal bait fishermen, Mida Creek Kenya, International Journal of Marine Science, 5(42): 1-9 (doi: 10.5376/ijms/2015.05.0042)
Marine overfishing is characterized by trophic cascades, due to selective removal of preferred top carnivores and confronts tropical fisheries managers in the absence of empirical evidence. This study compares the diets and trophic levels of fish landed by bait fishermen at Mida Creek, biosphere reserve, Kenya. Stomach content of fish landed using hook and line, baited with polychaete (choo) and hermit crab (dophe), and basket traps baited with mangrove whelk (tondo), were examined and compared. Results indicate that fish landed, consumed gastropod and fish, hence belong to 2.9th trophic level. Significant difference in trophic level occurred between gear and bait type. Hooks baited with dophe landed a variety of fish with a higher trophic level, such as Carangidae (TL>3.0), compared to other gears that landed lower trophic levels, such as Gerridae. Observed differences were attributed to gear and bait selectivity and differences in fishing grounds exploited. Absence of large top carnivores from landings is attributed to long-term overfishing pressure in nearshore fishing grounds. Additionally dietary overlap in mangrove dependent and creek fishes, landed by trap and hook fishermen, respectively, presents hitherto unexplored impacts.
Introduction
An indicator of global overfishing is the phenomenon of fishing down the food web that accompanies selective harvesting of large top predators from marine habitats (Duarte & Garcia, 2004; Cheunpagdee et al., 2006). Decline in trophic levels of exploited marine fish have raised concern over the sustainability of the fishery (Garrison & Link, 2000; Stergiou, 2005). Characterization of fish exploited, is crucial to understand both the status of a fishery, as well as the structure and functioning of the ecosystem, in order to drive sustainable utilization and management. Predation driven- top-down, planktivore driven- bottom-up and also intermediate trophic level driven- ‘wasp-waist’ systems, have been described (e.g. Duarte & Garcia, 2004; Buchheister, 2013).
Manifestation of fishing down the food web is marked by the disappearance of the apex predatory species from catches landed (Carpenter et al., 1985). Heavily exploited stocks report predominance of lower trophic levels and declining reproductive output, which precipitates trophic cascades (Garrison & Link, 2000).
Fishermen react to these changes by either exiting the fisheries, or changing fishing patterns. Among artisanal fishermen, wholly dependent on the catch for sustenance, exiting a fishery is rarely an option (Cinner et al., 2008). Changes in fishing patterns among artisanal fishermen involve alteration in gear use (e.g. using illegal gears), targeting previously unexploited stock (e.g. juveniles) and areas (e.g. encroaching on protected areas), that have negative impacts on fisheries and livelihoods.
The diet and feeding habits of fish is influenced by age, season, taxonomy, latitude, among others (Lopez-Peralta, 2002; Hammerschlag et al, 2010; Buchheister, 2013). Ontogenic shifts in fish diet have been demonstrated, while broadening of diet breadth and niche, occurs due to variation in food availability (Grutter, 2000; Smith et al., 2011; Sanchez-Hernandez et al, 2013). Marine fish belong to 2nd to 5th trophic levels, with ideal commercial species being piscivores, belonging to 3.5 to 4.5th trophic level (Sara & Sara, 2007). Tropical artisanal fisheries targeting dermersal fishes, such as Serranidae, Lutjanidae, belong to benthopelagic guilds at the 3 to 4th trophic levels, while flatfish (Dasyatidae) and sharks (Carcharhinidae), belong to the top carnivore levels (Kulbicki et al., 2005; Ainsworth et al., 2009). Trophic level monitoring is thus an important indicator of the sustainability of a multispecies artisanal fishery.
Tropical artisanal bait fishermen, employ basket traps and hook and line, baited with a variety of bait types, to catch fish in inshore waters (e.g. Glaesel, 1997; Elders of Atafu Atoll, 2012; Gomes, 2012). There is a paucity of published work on the diet and trophic interaction of fish landed by artisanal bait fishermen, especially in the Western Indian Ocean region. This study will evaluate the diet and trophic status of fish landed by bait fishermen using three types of bait at Mida creek, Kenya.
Materials and Methods
Study area
Mida Creek (03o 21’S; 39o 59’E) is a Marine National and Biosphere Reserve situated 88km North of Mombasa town, that has been in existence since 1968 (Figure 1). The creek covers an area of 31.6 km2, consisting of mangrove forests, seagrass beds, sandflats, rocky outcrops and subtidal habitats (Gajdzik et al., 2014).
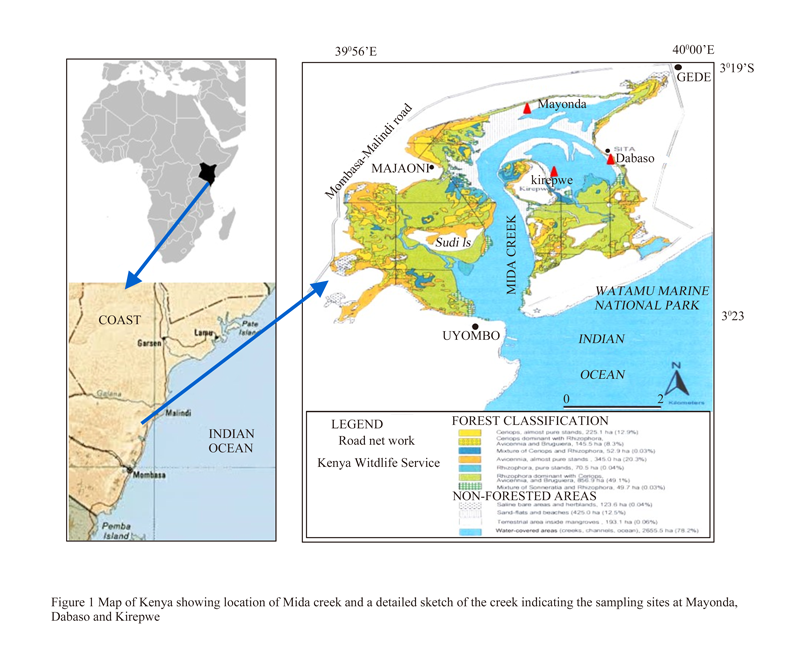 Figure 1 Map of Kenya showing location of Mida creek and a detailed sketch of the creek indicating the sampling sites at Mayonda, Dabaso and Kirepwe |
The communities living around the creek, estimated at 10,000 persons, are actively involved subsistence farming, small scale businesses and in fishing, employment in the tourism industry (Mwaipopo et al, 2011). The 2012 fisheries frame survey estimates between 250 and 500 registered fishermen, dependent on the Mida creek ecosystem (Frame survey, 2012). The Mijikenda (Kauma, Giriama, Chonyi, Jibao, Kambe, Ribe, Rabai, Duruma and Digo) and Bajuni dominate the fisher Community (Hoorweg et al., 2009). Additionally, migrant seasonal fishermen from Pemba (Tanzania) also participate in fisheries at Mida, during the Northeast monsoon period (October to January).
Trap fishing at Mida; The hexagonal basket traps used (2 x 3 m at widest part x0.5 m height), are woven using local fibers, and have funnel mouth opening of 6-8 cm and mesh openings of 3-5 cm on the sides. The traps are baited using crushed tondo (Terebralia palustris- Gastropoda; mangrove whelk), collected from the adjacent mangrove forest floor. Although other bait types are used elsewhere by trap fishermen, at Mida tondo are preferred (Kihia et al., 2013). Fishermen gather between 200 to 300 tondo, from within the mangrove forest. Trap fishermen insert 50 to 100 gastropods within the traps. Each trap fishermen owns and operates between three and five traps. The traps are ballasted with coral rock and retrieved after overnight soaking, within creeks in the mangrove forest.
Hook fishing at Mida; Bait used by hook fishermen is either choo (Marphysa sp- polychaeta) or dophe (Clibenarius sp-crustacea; hermit crab) gathered and prepared prior to the fishing. Although other bait types may be used at Mida, and elsewhere, hook fishermen at Mida prefer choo and dophe (Kihia et al., 2013). The local names of the bait i.e. choo- Marphysa and dophe- Clibenarius, will be used interchangeably in subsequent discussions. The choo are extracted at the seaward edge of the mangrove forest by digging and extraction from the sandy-muddy substrate and placed in suitable containers for subsequent fishing. On the other hand, the dophe are gathered by searching within the mangrove forest floor and collected for processing. The hermit crab shells are crashed and the crab within removed. The abdomen of the crab is cut off and subsequently used as the bait for fishing.
Hook fishermen attach appropriate bait to one or more hooks of size 8 to 18, and to a line, sinker and Styrofoam float. Bait used during hook and line fishing is carried in plastic containers and pieces of bait attached to the hooks. Hook fishing occurs from a dugout canoe and the line is reeled in by hand, and catch placed at the bottom of the canoe.
Fish sample Collection
Fish specimens were collected from fishermen during two sampling occasion in November, 2013 and February, 2014. Participating fishermen were identified from different fishing villages at Mida: Kirepwe, Mayonda and Dabaso, and monitored daily over a period of 10 days during each sampling occasion. The types of bait used as well as the fish landed were monitored. The fish landed were appropriately labeled and immediately frozen. In the laboratory, preserved specimen were defrozen and identified to species level using Fishbase (2011) and Richmond (2011) and stomach content removed and preserved in 5% formalin for further analysis.
Stomach content analysis
Dissected stomach content was estimated as empty stomach, quarter full, half full, ¾ full and full and allocated Hyslop Five point scale (0, 5, 10, 15 & 20 respectively). Food items in each stomach were sorted into major taxonomic groups under binocular microscope. The relative quantity of each food item was estimated, scored on the 5 point scale and recorded. Food index score was calculated using the following formulae; (Hyslop, 1980).
.png)
The trophic level (TL) of each fish was calculated using the following formulae as used by Choi et al, (2008);
![]()
Where TL- trophic level of fish species I, Pi- relative abundance of diet item i, Ti- trophic level of diet item i. Trophic levels of dietary items were obtained from Pauly and Froese (2000) as plant- 1, detritus- 1.5, finfish- 3.0, crab - 2.1, gastropod- 1.5, crustacean- 2.1. Data on diet and trophic levels obtained were compared among gear and bait used using Kruskal-Wallis ANOVA. What statistic used?
Results
A total of One hundred and twelve fish specimen from 10 families and belonging to 18 species were examined. Lethrinus harak (25%) and Gerres filamentosus (12%) dominated catches (Table 1). The next most important species formed between 5 and 10% of the catch and included; gerrids, lutjanids, lethrinids and carangids. The stomach content of fish landed by fishermen at Mida contained 6 main recognizable food categories; crabs, gastropod, fish, algae, shrimp and detritus. Average stomach fullness was estimated at 2.68±0.3 points. These stomachs were dominated by gastropod and fish that formed over 33 and 21%, of stomach content, respectively. Shrimp and algae each constituted less than 10% of stomach contents.
.jpg)
No significant difference in the dietary contribution of the food items between trap (2.65±0.5) and hook (2.74±0.2) users were detected (χ2=0.37, P>0.05). However, the stomach content of fish landed by trap fishermen was dominated by gastropod (8.38±1.2), fish (4.0±1.2) and contained low quantities of algae, shrimp and detritus. On the other hand, stomachs of fish landed using hooks contained all the 6 categories of food dominated by gastropod (4.60±0.4), fish (3.42±0.4) and detritus (3.06±0.4) were the next most important components, while shrimp and algae were least common (Figure 2).
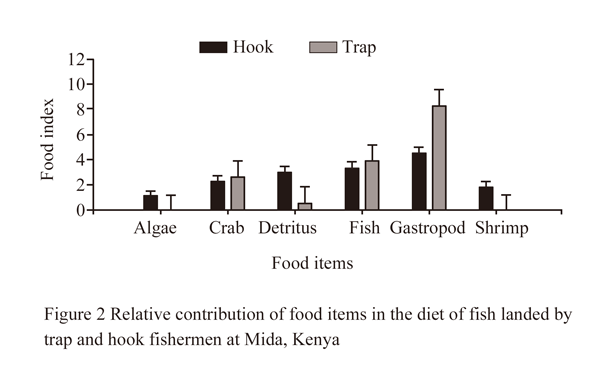 Figure 2 Relative contribution of food items in the diet of fish landed by trap and hook fishermen at Mida, Kenya |
The average trophic level of fish landed by bait fishermen at Mida was 2.88±0.06. There were significant differences in the trophic levels of fish landed by different gears (χ2=56.7, DF=28, P<0.001). Trap fishermen (2.76±0.1) recorded lower trophic level of landings than hook fishermen (2.91±0.1).
There were significant differences in the trophic levels of fishes landed by bait fishermen (χ2=56.7, DF=28, P<0.001). The highest tropic level species landed were secondary carnivores (TL<3.0) such as Trachinotes baiboni (3.10±0.5), Caranx ignobilis (3.4±0.1), Epinephelus malabaricus (3.34±0.5), Drepane punctata (3.5±0.5) and Terapon jarbua (3.5±0.5). While Gerres oyena (2.29±0.2) recorded the lowest trophic level (Table 1). Among the 17 species landed 12 belonged to the primary carnivore level, with a trophic level of between 2 and 3. Carangidae dominate the higher trophic level (2 species), while Lethrinidae (3 species), Gerridae (3 species) and Lutjanidae (2 species) dominate the primary carnivore guilds.
Hook gear users landed all the 18 types of fish, while the trap fishermen only landed 4 types of fish. Among the hook users, the highest trophic level fish landed was 3.8 for E. malabaricus, while the lowest was 2.4 for Gerres oyena (Figure 3). Among the trap fishermen, the highest tropic level species landed was Lutjanus fulviflamen (3.1), while the lowest was Gerres oyena (2.2).
There were no significant (χ2=4.46, P=0.6) difference in the dietary composition of fish landed using the three types of bait (2.60±0.3, 2.83±0.2, 2.76±0.5 for choo, dophe and tondo, respectively). However, there was significant interaction between bait type and food types. Stomach content of fish landed using choo bait was dominated by gastropods, fish and detritus (Figure 4). The stomach of fish landed with dophe was dominated by gastropod, fish and crabs, while the stomach of fish landed with tondo were dominated by gastropods. Additionally, fish landed with choo and dophe contained all 6 categories of food items, while those landed with tondo had only four items (crabs, detritus, fish and gastropods).
|
Figure 4 Composition of different food items in fish landed using different types of bait at Mida creek Discussion |
There were significant difference in the trophic levels of fishes landed by bait fishermen at Mida (χ2=91.58, DF=56, P<0.002). The highest trophic level fishes were landed by dophe fishermen (3.05 ±0.1), compared to lower levels by tondo-trap fishermen (2.74±0.1) and choo hook fishermen (2.72±0.1).
The composition and trophic level of fish landed differed with bait type. Dophe fishermen landed 14 types of fish, choo fishermen landed 11 types, while tondo fishermen landed 4 types of fish (Figure 5). Among dophe fishermen, the highest trophic level landed was Drepane punctata (3.81±0.2), while the lowest was Lutjanus argentimacus (2.44±0.3). Among choo fishermen, the highest trophic level species were Terapon jarbua (3.5) and Caranx ignobilis, (3.44), while the lowest was 1.9±0.4 for Lutjanus fulviflamen. Among tondo fishermen, the highest trophic level fish was L. fulviflamen (3.13±0.3), while the lowest was Gerres oyena (2.21±0.2).
.png) Figure 5 Trophic levels of fish landed using different bait a) choo, b) dophe and c) tondo by fishermen at Mida creek, Kenya |
Discussion
This study has demonstrated that Lethrinidae, Lutjanidae and Carangidae dominate landings at Mida with Lethrinus harak and Gerres filamentosus being the most common. It is apparent that artisanal bait fishermen at Mida exploit similar fish, as others in the region and Indo-West pacific region, where catches are dominated by fish from four main families; Lethrinidae, Serranidae, Lutjanidae and Carangidae (Kulbicki et al., 2005; Agembe et al., 2010). Agembe et al., (2010) report prevalence of Serranids in landings along the Kenyan coastline, thus the apparent decline of landings of the commercially important Serranid-Siganus sutor from Mida, is indicative of selective overharvesting, and may necessitate more exhaustive survey and subsequent regulation.
Fish landed by bait fishermen at Mida consumed crustaceans, molluscs, nekton, algae and detritus. Mollusc and finfish were the most important dietary component forming over 50% of stomach contents. Other dietary components, such as algae and detritus, formed less than 20% of items consumed. Hence fish landed by bait fishermen were generally carnivorous and thus their attraction to bait of animal origin. Predominance of gastropod in stomachs also suggests benthopelagic diet, as reported by Sara and Sara (2007). However, stomach fullness with gastropod and detritus items may relate to relative refractive nature of gastropod shell and plant matter. Softer bodied items, such as fish and also worms (polychaete and others) are easier to digest and hence have high gut clearance rates (Lopez-Peralta & Arcila, 2002). The absence of pure herbivores among fish landed may reflect the suitability of bait in targeting higher trophic level species. It is apparent that bait fishermen may only introduce top-down or wasp-waist impacts on a fishery in contrast to bottom up control where herbivores, dominate landings.
Fish landed by trap fishermen specialize on a diet composed mostly of gastropods but also fish, while that landed using hooks is more generalized, dominated by gastropod, fish and detritus. This is probably attributed to the trap fishermen preference for fishing within the mangrove forest where shading may preclude growth of algae and visual predators. Mangrove detritus is abundant within the mangrove creeks exploited by trap fishermen, however the recalcitrant nature of mangrove detritus and also high tannin content (Faraco & Lana, 2004; Kihia et al, 2011), may limit their usefulness as food for fish. Similarly, (Heminga et al., 1994) reported minimal mangrove derived signatures in fish captured in seagrass beds adjacent to mangrove forests. On the other hand, broadening of the food resource base by fish in the open creek, that are landed by hook fishermen, may occur due to the lower productivity of the nutrient deficient open sea compared with the mangrove forest. The contribution of herbivorous mangrove benthic invertebrate detritivores to nutrient cycling appears to support the fisheries and require further study. Bait overharvesting from within mangrove habitats, may however impact benthic community structure, with consequences on fishery.
Dietary composition was dependent on bait used, with fish landed using tondo being more specialized on gastropods, while those landed using dophe and choo were generalist with most food components involved. Difference between specialist diet fish caught by tondo users and the more generalist diet of fish landed using choo and dophe may also be attributed to the different habitats exploited for fishing. The tondo fishermen exploit fishing grounds within creeks and rivulets in the mangrove forest. Migrant Pemba fishermen however have skills enabling them to exploit the less fished open sea grounds using larger traps compared to the smaller traps of the local Mida fishermen. Hence use of alternative open sea fishing grounds may provide respite for overexploited mangrove dependent fishes. Fish landed using choo consume gastropod, fish and detritus, while dophe fishermen land fish that consume gastropod, fish and crabs. This may indicate difference in attractiveness of bait used by hook fishermen.
Bait fishermen at Mida, landed fish with an average trophic level of 2.9. This indicates that the fish feed above the carnivore guilds. In marine habitats, this corresponds to midlevel trophic levels as opposed to the 4th or 5th top carnivore levels. This may indicate fishing down the food web as a result of overfishing pressure. Pasquaud et al (2010) reported similar range of trophic levels (2.8-3.9) from an estuarine fishery as reported here. However, McClanahan and Mangi (2004) reported hook fishermen as landing higher trophic level fishes (3.6) compared to basket trap (2.5) and other gears (> 2.5). Decline in actual trophic levels from those previously reported by McClanahan & Mangi (2004), may thus indicate overfishing since the last survey. This is especially prevalent among hook fishermen where fisher landing trophic level seem to have declined from secondary carnivore to primary carnivore guilds.
Lower trophic level reported here may also be due to lack of requisite capacity, such as gears and vessels, among Mida fishermen that limit them to shallow nearshore mangrove dominated grounds, where the larger top carnivores, such as sharks, barracuda, tuna and others, are less prevalent. However, occasional landings of top carnivore, such as rays at Mayonda, were observed, but these specialized fishermen used shrimps and small fish as bait, and were therefore not included in the current study. It is possible that improvement in gear and skills may emancipate fishermen from overfishing within mangrove and near shore grounds, and enable forays into deeper and more distant open water grounds, where fishing may be more profitable to livelihoods. Similarly exploitation of alternate bait may also affect targeted fish.
Fishermen using tondo baited traps and hooks baited with choo, land lower trophic level fish than their dophe hook counterparts. Observed differences between tondo and dophe fishermen are probably related to the locality in which these gears are active. Most trap fishermen placed their traps within mangrove creeks, and hence may be targeting species that either consumes detritus or that feed on herbivorous mangrove fauna dependent directly on plant matter, such as crustaceans. Epibenthic megafauna, such as grapsid and sersamid crabs, as well as mangrove whelks, dominate and are mangrove keystone species, that influence mangrove litter processing. However, difference between choo and dophe fishermen may reflect difference in attractiveness of bait to fish with dophe attracting higher carnivores than choo fishermen.
Over 75% of fish landed by the bait fishermen belonged to above the 3rd trophic level primary carnivore guild. The carangidae were the highest trophic level fish landed by fishermen at Mida, while the lowest were Gerridae. Difference in feeding habits among these fishes is attributed to taxonomic difference coupled with size differences. Larger fish consume more nekton and mollusc than smaller fish and juveniles (Kulbicki et al., 2005). Shading provided by overhead mangrove trees protects juvenile fish from visually searching pelagic predators. Thus benthic foraging may be a more fruitful lifestyle. Baited hook fishermen land six times greater fish variety than trap users. Among dophe-hook users, 5 fish type were landed (e.g. Epinephelus, Caranx, Drepane, Terapon) belonging to above the 3rd trophic level, compared to 3 among choo users and only Lutjanus among trap users.
In addition, different trophic levels were recorded for the same fish type among the gears. Lower trophic level was reported in trap fisher landing compared to hook fisher for Gerres oyena and G. longilostris and Epinephelus (3.81 & 2.9 for hook and trap, respectively). In contrast, Lutjanus fulviflamen landed by hook fishermen was of a lower trophic level than that landed by trap fishermen (2.58 & 3.13 for hook and trap, respectively). However, Lethrinidae trophic level remained similar for both traps and hook fishermen (2.84 & 2.97 for hook and trap, respectively). These variations may be attributed to change in fish diet due to ontogenic shift at the two habitats (Gullström et al, 2012), exploited by trap and hook fishermen. Generally, within the highly turbid mangrove creek (where trap fisher place traps), consumption of detritus and other dead organic matter may contribute to much of the fish diet, as compared to visual predation on mobile higher trophic level prey items in the open sea (where hook fishermen ply). School forming fish, such as the Lethrinidae may retain their ability to hunt for groups of juvenile fish prey, hiding within mangrove creeks and hence their ability to maintain diet in both fishing grounds.
Fishermen using the crustacean dophe bait, targeted and landed higher trophic level fish than fishermen using either mollusc (tondo) or polychaetes (choo). In addition, dophe fishermen landed a higher variety of fish than tondo fishermen. This indicates that dophe is a generalist bait, attracting higher carnivores as compared to either choo or tondo.
Dophe fishermen landed 6 types of fish (Drepane, Coris, Plectorinchus, Terapon, Scarus, Trachinotes), that were not landed by any of the other bait types. On the other hand, choo fishermen landed 3 fish types (G. longilostris, Gobius keiensis and L. nebulosus) that were unique. Epinephelus was only landed by trap and dophe fishermen, while Gerres oyena was only landed by choo and tondo fishermen. Since there were no reported alterations in fishing ground exploited by fishermen examined during this study, these difference may indicate variations in attractiveness to bait to the fish targeted by the fishermen. Bait attractiveness functions through both visual and chemical signals that attract fish to bite or trapping zone.
In the highly turbid mangrove dominated creek, proteinious chemical cues emanating from the bait may play a powerful role in enhancing attractiveness of bait to fish. However, the chemical nature of the baits used by artisanal fishermen has rarely been explored. This also indicates that although choo and tondo fishermen attract similar trophic level fish, the variety landed by trap fishermen is lower. For instance, the mangrove snapper Lutjanus fulviflammen juveniles may use the open creek waters (landed by choo fishermen-TL=1.9) and move into mangrove habitats as adults (landed by tondo fishermen-TL=3.1). This requires further elaboration by examination of ontogenic shifts in species landed by bait fishermen.
Gajdzik et al., (2014) describe the trophic interaction of fish caught using nets at the same study site (i.e. Mida creek) reporting a total of 27 species comparable to total number of species landed by all the baited gears reported here. This coincides with the well documented fact, that bait fishing is more selective. However, such selectivity only applies when one or a few baited gears are used, but where a variety of bait and gears, are used the selectivity advantage of such gears is lost. Additionally, closure scrutiny of the species involved indicates significant differences among the gears, with net landings dominated by transient juvenile piscivorous barracuda (Syphaerena) and zoobenthivorous Gerridae. Most species landed using nets are thus apparently different from those landed using bait as reported here. These differences may be attributed to net fishermen targeting shoaling pelagic fishes, while the bait fishermen target dermersal species. The use of small mesh size nets is frequently stringently controlled in order to increase selectivity of these gears. However, the use of gears baited with tondo and choo as reported here, target lower trophic level fish compared to dophe bait. In addition, the use of small hooks (>#14) may also increase proportion of juveniles landed and requires further elaboration.
Conclusion
Fishermen at Mida use hooks and traps baited with choo, dophe and tondo to target mid-level carnivores at 2.9th trophic level. Gear and bait types have significant influence on diet and trophic status of fish targeted. Hook fishermen using dophe landed a larger variety of generalist and higher trophic level fishes, compared with trap fishermen that used tondo bait that targeted specialist species of lower trophic levels. The adoption of a variety of gears and baits by artisanal fishermen, targeting a mixed range of fish with broad niche requirements, may spread the risk of trophic cascades that accompany overfishing. Interventions that seek to narrow bait and gear variety in the absence of effective effort control may thus be inappropriate in maintaining artisanal fisheries integrity and sustainability.
Acknowledgement
Research grant was provided by the National Commision of Science and Technology, Kenya, awarded to A Muthumbi, CM Kihia & J Okondo. Institutional support and laboratory facilities provided by Nairobi and Egerton Universities, Kenya Marine and Fisheries Institute are appreciated. Members of the Mida Creek Conservation Community, especially J. Mwamure, assisted in introduction to fishermen. This paper formed part of Bsc thesis by Hendrick Y. (Egerton University), while A Nthiga and V Njuguna (Msc Students University of Nairobi students) participated in both field and laboratory analysis. J. Ngatia, S. Muhamed and M. Abutrika (University of Nairobi Bsc students), participated in fieldwork and also B. Otieno (KMFRI), assisted in identification of fish. The contributions and suggestions made by the anonymous reviewers are also appreciated.
References
Agembe S, Mlewa C.M., and Kaunda-Arara B, 2010. Catch composition, abundance and length-weight relationship of groupers (Pisces: Serranidae) from inshore waters of Kenya. Western Indian Ocean Journal of Marine sciences, 9(1): 91-102.
Ainsworth C.A., Kaplan I.C., Levin P.S., and Mangel M., 2009. Diet composition of fish species in North Gulf of California. Hem Nalini Morzaria Luna (Ed), A handout of the integrated Marine Ecology. Ecosystem Science Program, Northwestern Fisheries Science Centre, NOAA, Seattle USA.
Buchheister A., 2013. Structure, drivers and trophic interaction of the dermersal fish community in Chesapeake Bay. PhD thesis Faculty of Marine Science, College of William and Mary in Virginia, USA.
Carpenter S.R., Kitchell J.F., and Hodgson J.R., 1985. Cascading trophic interactions and lake productivity. Bioscience, 35(10): 634-639
http://dx.doi.org/10.2307/1309989
Cheunpagdee R., Liguori L., Palomares M.L.D., and Pauly D., 2006. Bottom-up, global estimates of small scale marine fisheries catch. Fisheries Centre Research Report Vol 14(8).
Choi Y.M., Yoo J.T., Choi J.H., Choi K.H., Kim J.K., Kim Y.S. Kim and Kim J.B., 2008. Ecosystem structure and trophic level to the oceanographic conditions around the waters of Jeju Island. Journal of Environmental Biology, 29(4): 419-425.
Cinner J.E., Daw T., and McClanahan T.R., 2008. Socioeconomic factors that affect artisanal fishers readiness to exit a declining fishery. Conservation Biology 23(1): 124-130
http://dx.doi.org/10.1111/j.1523-1739.2008.01041.x
Duarte L.O., andGarcia C.B., 2004. Trophic role of small pelagic fishes in a tropical upwelling system. Ecological Modeling, 172: 323-338
http://dx.doi.org/10.1016/j.ecolmodel.2003.09.014
Elders of Atafu Atoll, 2012. Echoes at fisherman’s rock; Traditional Tokelau fishing. Eds A Hooper & I Tinielu, Knowledge of Nature, 4, UNESCO, Paris, 120pp.
Faraco L., Ditze F., and Lana da Cunha P., 2004. Leaf consumption levels in subtropical mangrove of Paranagua Bay (SE Brazil). Wetlands Ecology and Management, 12: 115-122
http://dx.doi.org/10.1023/B:WETL.0000021666.42546.c2
Fishbase 2011. Fishbase 2011. Froese JE and Pauly D (Eds) www electronic publications.
Frame survey, 2012. Marine fisheries frame survey. Ministry of fisheries Development, Fisheries Department, Nairobi, Kenya, 85pp.
Gajdzik L., Vanreusel A., Koedam N., Reubens J., and Muthumbi A.W.N., 2014. The mangrove forest as a nursery habitat for the ichthyofauna of Mida creek (Kenya, East Africa). Journal of the Marine Association of United Kingdom, 1-13
Gallström M., Dorenbosch M., Lugendo BR.., Mwandya A.W., Mgaya Y.D., and Berkström C., 2012. Biological connectivity and nursery function of shallow ^water habitats in Chwaka bay. In Torre-Castro M., T.J Lyimo (Eds) People Nature and research in Chwaka Bay, Zanzibar, Tanzania, 175-192 WIOMSA, Zanzibar.
Garrison L.P., and Link JS, 2000. Fishing effects on spatial distribution and trophic guild structure of the fish community in the Georges Bank region. ICES journal of Marine Science, 57: 723-730
http://dx.doi.org/10.1006/jmsc.2000.0713
Glaesel H., 1997. They are not just ‘samaki’: towards an understanding of fisher voices on the Kenyan coast. AAP, 51: 165-179
Gomes I., 2012. Artisanal fisheries analysis within the Mpunguti marine reserve (Southern Kenya): Gear based management towards sustainable management. Msc thesis, Erasmus Mundus Programme, Algarve University, Spain.
Grutter A.S., 2000. Ontogenic variation in the diet of the cleaner fish Labroides demidiatus and its ecological consequences. Marine Ecology Progress Series, 197: 241-246
http://dx.doi.org/10.3354/meps197241
Hammerschlag N., Ovando D., and Serafy J.E, 2010. Seasonal diet and feeding habits of juvenile fishes foraging along a subtropical marine ecotone. Aquatic Biology, 9: 279-290
http://dx.doi.org/10.3354/ab00251
Hemminga M.A., Slim F.J., Kazungu J., Gansse G., Nieuwenhuize J., and Kuyt M.M., 1994. Carbon outwelling from a mangrove forest with adjacent seagrass beds and coral reefs, Gazi bay, Kenya. Marine Ecology Progress Series, 106: 290-301
http://dx.doi.org/10.3354/meps106291
Hoorweg J., Wangila B., and Degan A, 2009. Artisanal fisheries on the Kenyan coast; household livelihood and Marine Resources Resource Management. Leiden 146pp
Hyslop E.J., 1980. Stomach content analysis: A review of methods and their application. Journal of Fishery Biology, 17; 411-429
http://dx.doi.org/10.1111/j.1095-8649.1980.tb02775.x
Kihia C.M., Mathooko J.M., Ruwa R.K., and Shivoga W.A., 2011. Influence of human disturbance on leaf herbivory at the Gazi mangrove forest, Kenya. African Journal of Aquatic Sciences, 30(3): 1-7
http://dx.doi.org/10.2989/16085914.2011.636901
Kihia C,M,, Muthumbi A., andOkondo J., 2013. Fisheries bait harvesting among artisanal fishers at Mida creek, Kenya. Proceedings of the 8th WIOMSA Symposium, Maputo, Mozambique.
Kulbicki M., Bozec Y., Labrose P., Letourneur Y., Mao-Than G., and Wantiez L, 2005. Diet composition of carnivorous fishes from coral reef lagoon of New Caledonia. Aquatic Living Resource, 18: 231-250
http://dx.doi.org/10.1051/alr:2005029
Lopez-Peralta R.H., and Arcila C.A.T., 2002. Diet composition of fish species from the Southern Continental shelf of Columbia. Naga World Fish Quarterly, 25 (3&4): 23-40
McClanahan T.R. and Mangi S.C., 2004. Gear based management of a tropical artisanal fishery based on species selectivity and capture size. Fisheries Management and Ecology, 11: 51-60
http://dx.doi.org/10.1111/j.1365-2400.2004.00358.x
Mwaipopo R., Fischer E., Wanyonyi I., Kimani P., Tunje J., Msuya F., and Bashemerewa V., 2011. The relationship between community based organizations and the effective management of coastal and marine resources in the WIO region. A report for the WIOMSA, xii; 85pp.
Pasquaud S., Pillet M., David V., Salitour B., and Elie P., 2010. Determination of fish trophic level in an estuarine system. Estuary and Coastal Shelf Science, 86: 237-246
http://dx.doi.org/10.1016/j.ecss.2009.11.019
Richmond M.D., 2011. A field guide to the seashores of Eastern Africa and the Western Indian Ocean Islands. 3rd Ed, Sida/WIOMSA; 464pp
Sanchez-Hernandez J., Servia M.J., Vieira-Lanero R, and Cobo F., 2013. Ontogenic dietary shift in predatory fish species: The brown trout as an example.
http://dxdoi.org/10.5772/54133
Sara G., and Sara R., 2007. Feeding habits and trophic level of blue fin tuna Thunnus thynnus of different size classes in Mediterranean Sea. Journal of applied Ichthyology, 23(2): 122-127
http://dx.doi.org/10.1111/j.1439-0426.2006.00829.x
Smith J.A., Baumgartner L.J., Suthers I.M., and Taylor M.D., 2011. Generalist niche, specialist strategy: the diet of an Australian percichthryid. Journal of fish Biology, 78: 1183-1199
http://dx.doi.org/10.1111/j.1095-8649.2011.02926.x
Stergiou K.I., 2005. Fisheries impact on trophic level: long term trends in Hellenic waters. In Papathanasious E & A Zenetos (Eds), State of the Hellenic Marine Environment, Hellenic Centre for Marine Research, Athens, Greece, pp326-329



. PDF(717KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Kihia C.M.
. Hendrick Y.
. Muthumbi A.
. Okondo J.
. Nthiga A.
. Njuguna V.M
Related articles
. Bait fishermen
. Polychaete
. Hermit crab
. Gastropod
. Hook
. Traps
. Trophic levels
Tools
. Email to a friend
. Post a comment