Decoupled Seasonal Stress as an Indication of Chronic Stress in Montastraea cavernosa and Porites astreoides Inhabiting the Florida Reef Tract 
2 Annis Water Resources Institute, Grand Valley State University, Muskegon, Michigan, United States of America
3 Woods Hole Oceanographic Institution, Woods Hole, Maine, United States of America
 Author
Author  Correspondence author
Correspondence author
International Journal of Marine Science, 2016, Vol. 6, No. 21 doi: 10.5376/ijms.2016.06.0021
Received: 21 Jan., 2016 Accepted: 25 May, 2016 Published: 25 May, 2016
Haslund J.A., Hauff B., Strychar K.B., and Cervino J.M., 2016, Decoupled seasonal stress as an indication of chronic stress and site dependent responses in Montastraea cavernosa and Porites astreoides inhabiting the Florida Reef Tract, International Journal of Marine Science, 6(21): 1-20 (doi:10.5376/ijms.2016.06.0021)
Although mass mortality from disease and climate anomalies are largely to blame for rapid losses in coral cover along the Florida reef tract, the failure of extant populations to recolonize the offshore zone is puzzling given improvements to water quality and narrower range in seawater temperature compared to the inshore patch reef zone. Using existing data from the Coral Reef Environmental Monitoring Program (CREMP) and from an inshore and offshore site established to compliment this dataset we identified significantly increased abundance and diversity of scleractinian coral from inshore to offshore across the Florida reef tract. Applying exploratory statistical methods we identified two abundant species, Montastraea cavernosa and Porites astreoides, inhabiting both zones to varying degrees. Following reciprocal transplantation of conspecifics between a representative offshore and inshore reef (6 m depth), we monitored monthly coral colony brightness (a measurement related to endosymbiotic dinoflagellate density) over a two-year period to examine symbiont loss, a common stress response in scleractinian corals. Although species-specific stress patterns were not identified, zone-specific variation was evident. Trigonometric regression of stress level by month revealed a significant relationship supporting an annual stress and recovery period at the inshore patch reef zone. Contrary to this result, conspecifics transplanted to the offshore zone did not display recovery resulting in a continued chronically bleached state over the two-year period. Our results implicate alleviation or decreased stress at inshore sites and the importance of extending greater protection to reefs within this zone.
Introduction
The Florida reef tract has experienced dramatic decreases in coral cover (Aronson and Precht, 2001), shifts in reef communities (Pandolfi et al., 2005; Ruzicka et al., 2013), and decreases in reef architectural complexity (Alvarerz-Filip et al., 2009), which have all resulted in decreased biodiversity. This trend is most apparent along the 15 offshore reef preservation areas of the Florida Keys National Marine Sanctuary (FKNMS). Located between 1 and 8 m depths and 5 to 7 km offshore, these once highly productive reef crest and fore reef ecosystems no longer support accreting scleractinian coral communities and can be considered relict in comparison to historical baselines (Palandro et al., 2008). This imperiled status of the offshore reef zone is not a result of a single stressor. Diseases and macro-algal overgrowth arising from human interaction, decreased coral resilience following destructive hurricanes and anomalous hypothermic (Hudson et al., 1976; Walker et al., 1982; Shinn et al., 2003) and hyperthermic seawater temperature events have all greatly affected this region. Presently, conditions are such at offshore reefs that corals should display net accretion, however reef building coral populations remain at low abundances. Observing transects extending from inshore to offshore, coral abundance decreases to the offshore bank reef zone (Roberts et al., 1982; Lirman and Fong, 2007; Schutte et al., 2010) with approximately two-times lower abundance at offshore reefs. In contrast, sustained growth has been observed throughout the inshore patch reef system despite far fewer protected areas in contrast to offshore reefs, increased frequency of hyper and hypothermic seawater temperature events experienced (Soto et al., 2011), increased dissolved inorganic nitrogen, soluble reactive phosphorus, and chlorophyll a, increased turbidity, greater macro algal biomass (Lapointe et al., 2004), and closer proximity to anthropogenic sources of stress (Ginsburg et al., 2001). Several hypotheses have been proposed regarding the proliferation of inshore reefs and the deterioration of offshore reefs including (1) continued persistence of large fecund colonies in inshore reef zones resulting in increased inshore recruitment, (2) diet supplementation from increased turbidity and nutrient concentration as a means to alleviate stressful near-shore conditions, and (3) differences in seawater temperature variance resulting in a wider temperature range allowing corals inhabiting inshore reefs the opportunity to acclimate to future changes in seawater temperature (Soto et al., 2011).
Recruitment does not appear to be responsible for the differences in coral abundance between adjacent inshore and offshore sites. The current state of knowledge emphasizes the importance of local recruitment for the maintenance of coral population abundance (Mumby et al., 2008; Steneck et al., 2009) and low frequency immigrant settlement promotes genetic diversity (Sammarco et al., 1989; Noreen et al., 2009). Co-implementation of larval dispersal models, genetic population connectivity (Foster et al., 2012) and sensitive identification and quantification of recruits (Schmidt-Roach et al., 2008; Hsu et al., 2014) has increased the accuracy of coral recruitment estimation. In the Florida Keys annual recruitment at inshore and offshore reefs is highly variable (Moulding, 2005; Shearer and Coffroth, 2006) and not a locally defining characteristic (Miller et al., 2000). Post settlement processes and stress (i.e. hypotheses 2 and 3) are therefore more likely correlated to the difference in reef growth and community structure observed.
We hypothesize that corals inhabiting inshore reef zones are able to respond to a wider range of stress levels as a consequence of frequent occurrences of non-lethal stress levels characteristic of this zone. The history of abiotic and biotic stress experienced by a reef affects the inhabitants response, growth, selection for resistant individuals, and in the case of corals establishment of resistant symbionts following aberrant levels of stress (Haslun et al., 2011; Palumbi et al., 2014). Gradients of increased levels of abiotic factors associated with stress extend from offshore reefs to inshore reefs in the Florida Keys (Lapointe et al., 2004) and supporting this hypothesis a negative correlation exists between both coral cover and colony size, as well as distance from shore (Lirman and Fong, 2007). The stress level experienced by the organism is most likely not a reflection of any one stressor but rather the cumulative effect of stressors present. The level of one factor may provide a refuge from a factor the organism is more susceptible to, decreasing the severity of a potentially lethal stressor. For example, increases in chlorophyll a may occur concomitantly with increases in turbidity (Ziemann et al., 1992). Increased turbidity may in-turn, decrease photic stress on scleractinian corals during increased water temperatures resulting in an overall decreased level of stress.
Identification of abiotic factors contributing to the community level differences in abundance and size of corals inhabiting inshore and offshore reefs is critical to the management of this habitat (Lirman and Fong, 2007). This study integrated and applied coral cover data from the coral reef environmental monitoring project (CREMP), environmental data from the water quality monitoring project (WQMP), and coral community and environmental data from ongoing research projects coordinated by our lab group to describe the asymmetry of coral cover currently observed between the inshore patch reef system and offshore bank reef system. We identify four coral species with differential abundances within the nearshore and offshore zones, the study of which may increase our understanding of the juxtaposition in coral growth between these environments. To provide a more complete understanding of the community dynamics of a symbiotic organism we also observed the seasonal photosynthetic competence (symbiont photo-physiology) and colony color of the identified species at two sites, one representative of an inshore patch reef and the other representative of an offshore bank reef, over a two-year period. From this information the seasonal response strategy of each species was determined.
2 Materials and Methods
2.1 Coral Communities of the Patch Reef and Bank Reef Systems
Relative abundances of corals inhabiting offshore bank reefs and inshore patch reefs, spanning a 16-year period, were obtained from CREMP (http://ocean.floridamarine.org/FKNMS_WQPP/pages/cremp.html). This dataset has been applied to the detection of potential causal factor for regional differences in benthic communities coupled with the WQMP (http://ocean.floridamarine.org/FKNMS_WQPP/pages/wqmp.html) (Maliao et al., 2008). This previous work indicated that the CREMP monitoring strategy correlates well with the results of other sampling efforts including the Atlantic and Gulf Rapid Assessment protocol (AGRRA; http://www.agrra.org/). The CREMP and WQMP datasets (1997-2012) were used to establish the extent of variability in coral communities and local environment between the inshore and offshore zones and sites associated within each zone. Of the available reefs within the two datasets we selected reefs within the lower and middle keys for three reasons. Firstly, throughout the lower and middle keys regions there are a number of offshore bank reefs as well as patch reef formations that spatially parallel one another (Figure 1). Secondly, for the past three years we have monitored an inshore patch reef and an offshore bank reef at the heart of this distribution, Birthday and Acer 24 reefs respectively (Figure 1). Thirdly, this region has been characterized by decreasing gradients of nutrients, turbidity and temperature from the inshore to offshore zones. Differences in these factors among sites and between the inshore and offshore zones (i.e. nutrients, turbidity, and temperature) may provide a foundation to understand the evident variation in coral cover.
.png) Figure 1 Map of Lower and Middle Florida Keys Study Sites. Sites were either selected from the Coral Reef Environmental Monitoring Project (Diagonal Labels) or from sites established for this study (Horizontal Labels). Sites identified with open triangles indicate offshore sites (n=6) while those with open circles indicate inshore patch reef sites (n=6). |
Reefs with greater than 10% total coral cover as of 1996 were included. Reefs with cover less than 10% as of 1996 may have already become relict or may never have supported a large community of corals and were therefore excluded. From this criteria, five CREMP sites remained as representative of each of the two zones. Inshore sites included Western Head, Cliff Green, West Washerwoman, Dustan Rocks (also referred to as East Washerwoman), and Western Turtle Shoal. West Washerwoman is the largest patch reef of this set and is considered the largest in the lower keys. East Washerwoman and Western Turtle Shoal are also considerably large patch reef groups while Western Head and Cliff Green are singular patch reefs see (Lidz et al., 2007) for mapping and description of these patch reefs). Offshore reefs included Sand Key, Rock Key, Eastern Sambo, Looe Key, and Sombrero Key. Each of these offshore bank reefs is characterized by spur and groove formations preceded inshore by patch reefs.
2.2 Coral Community Characteristics at Inshore and Offshore Reefs
Colony size and abundance of corals were assessed biannually from 2011 to 2013 using photo-transects established at our inshore patch reef site (Birthday reef: 24.57917’ N, -81.49692’ W) and offshore bank reef site (Acer 24 reef: 24.55268’ N, -81.43741’ W), both located at depths of 6.5 m (Figure 1). Assessments were conducted by divers using self-contained underwater breathing apparatus (SCUBA) during winter (February) and summer (August) seasons. Divers photographed the benthos at random non-overlapping points (n = 15) along 30 m transects (n = 3) with a Nikon D5100 (Nikon Inc., Melville, NY) camera affixed to a 0.5 x 0.5 m quadrapod (Coyer et al., 1999). Lighting was supplied by a Fantasea NanoFlash strobe (Fantasea, Blaine, WA). Transects were measured using surveyor tape and extended out from a central location (the reciprocal transplant location at each reef) in three randomly assigned directions. Each direction was selected with a random number generator sampling points between 0 and 360 in increments of 5. The photo-quadrat method has been shown to be the most cost effective method to assess coral and sessile benthic communities without sacrificing accuracy (Leujak and Ormond, 2007). Images were analyzed using Coral Point Count with Excel (CPCe) extensions (Kohler and Gill, 2006). Coral species richness, abundance, area, percent cover, as well as benthos composition including coral, sponge, algae, sand, and rubble were determined. Further, all individual Montastraea cavernosa and Porites astreoides colonies within 1 m of transect were analyzed for photosynthetic capacity using a Diving Pulse Amplitude Flourometer (PAM: Walz). We were unable to assess field corals in a dark adapted state and therefore obtained the effective quantum yield of photochemical energy conversion (ΔF/Fmˊ). The change in F (ΔF) represents the change in fluorescence from the maximal fluorescent yield of the coral in an illuminated environment (Fmˊ) following a saturating pulse from the instrument. The use of these two measurements has been found to be highly correlate with quantum yield (Genty et al., 1989). The effective quantum yield was determined at the apex of colonies with the fiber quantum sensor held within the Surface Holder, to maintain a distance of 10 mm between it and the colony tissue (Cervino et al., 2012).
2.3 Location-Dependent Colony Brightness, a Bleaching Characteristic
Following the identification of species characteristic of the inshore patch reef and offshore bank reef systems, a reciprocal transplant was carried out between Birthday Reef and Acer 24 with two species of coral, M. cavernosa and P. astreoides. Coral fragments (n=10; 16 x 16 cm) of both species were collected from colonies at least 10 m apart at each reef using a steel mallet and cold chisel. Permission for field work at Birthday Reef and Acer 24 was granted by National Oceanic and Atmospheric Administration National Marine Sanctuaries (Permit # FKNMS-2011-107). Fragments were transferred in large coolers filled with site-derived water to the Mote Marine Laboratory Tropical Research Laboratory (MML), where they were immediately sectioned into two 8 x 8 cm fragments with a tile saw lubricated and cooled with sterile artificial sea water (Instant Ocean) sprayed on the blade with a wash bottle. Following sectioning, fragments were transferred to MML flow-through seawater raceways. Raceways were shaded from direct sunlight to decrease stress. Following 2 days of recovery, fragments were attached to pucks (1 part concrete: 3 parts aragonite sand) with a two-part epoxy (All Fix Epoxy; Philadelphia, PA USA). After another 3 days of recovery, corals were transferred in large coolers filled with seawater from the MML flow-through system, to each field site. To complete the reciprocal transplant, ½ of each colony was returned to the site of collection as a control and the other ½ transplanted to the alternate site. Each field site consisted of 6 concrete blocks affixed to the calcium carbonate substrate using a 1 part plaster: 2 parts concrete mixture in a hexagonal shape. Each concrete block consisted of 6-7 randomly assigned coral fragments of a single species. Fragments were attached to each block with All Fix Epoxy. Neighboring blocks harbored different species. We observed growth/ skeletal accretion in all coral fragments during the study, indicating that the sampling and transplant procedures employed did not greatly impact coral physiology.
Images of each cinder block and its associated coral fragments, were taken monthly over the course of two years with a Nikon D500 in a Fantasea underwater housing. Illumination was provided by the built-in camera flash and housing diffuser. Each image was analyzed with Image J to determine the mean fragment brightness. The brightness of gray-scale images has been shown to be correlated (R2 = 0.63) with symbiont density (Siebeck et al., 2006) and thus, colony darkening (decreased brightness) is an indication of increased concentration of the coral endosymbiont (Siebeck et al., 2006). All brightness values were determined relative to a clean area of the cinder block (the brightest object in each image). Analysis proceeded by first transforming an image to 8-bit format. The scale of each image was set using the average width of a cinder block (15.24 cm). Regions were drawn around each fragment including only live coral tissue using the polygon tool and minimum, maximum, and mean brightness values obtained. Differences in the background brightness between different blocks and/or site dependent water quality (turbidity) was corrected for by adding the corresponding value or subtracting the corresponding value to reach that of the first sampling period for each block. For analysis, mean colony brightness values were either used as is or grouped into one of three bins of equal width spanning the range of brightness values observed.
2.4 Statistical Analyses
Statistical analyses were performed with R version 3.0.2 (R Core Team, 2013) and all figures were created with ggplot (Wickham, 2009).
We first analyzed the CREMP dataset for variation in coral cover among sites and zones (inshore vs. offshore) across the 16-year sampling period via two-way repeated measures analysis of variance (ANOVA). The aov() function within the statistical package R was applied with reef zone and sites as predictors of total coral cover and an error term described by sample year to create a repeated measures design. Assumptions of ANOVA were visually evaluated with the plot() function, which produces figures including residuals vs. the fitted model, standardized residuals vs. theoretical quantiles (qq-plot), square root of the standardized residuals vs. the fitted values, and the standardized residuals at each factor level. Non-linearity in these figures is indicative of a deviation from a normal distribution. Homogeneity of variances was assessed using the Bartlett test (bartlett.test()), which tests the null hypothesis that variances are equal at all levels.
Site community richness and evenness was assessed across the 16-year CREMP dataset and the 2 year dataset from our field sites using Rényi diversity profiles (Jost, 2006). The renyi () function from the vegan package served as the platform for this analysis (Oksanen et al., 2013). Mean relative abundances of coral species across the 16-year period were applied to this function. The shape of each profile provides information with respect to the evenness of a community. Horizontal lines represent perfectly even communities where all species are present in equal abundances. Alpha values of 0, 1, 2, and infinity indicate species richness, Shannon diversity index, Simpson index (reciprocal), and the proportion of the most abundant species present, respectively.
Coral community differences were identified using a multivariate approach. Principal coordinate analysis (PCoA) was performed on the annual relative abundances of coral species at each site. To accomplish this, Bray-curtis dissimilarities were calculated with the vegdist () function from the vegan package (Oksanen et al., 2013). Eigen analysis was performed by the application of the cmdscale () function on this dissimilarity matrix in 2 dimensions to produce eigen values. Confidence intervals (95%) were calculated from these eigen values across years for each site. The correlations between the original relative abundances and eigen values were used as loadings to determine the association of species with particular sites.
Principal component analysis was carried out on abiotic factors including ammonium (NH4+) total organic nitrogen (TON), chlorophyll a (Chl a), total organic carbon (TOC), silicate (SiO2), salinity, SWT and dissolved oxygen (DO) from the WQMP. The WQMP sites mirrored the selected CREMP sites. Data were standardized by subtracting each measurement by the mean and dividing by the standard deviation. The covariance of this matrix was then calculated and eigen analysis performed on the covariance matrix with the eigen () command (Jackson, 1980). The first three principal component axis were visualized in biplots to display how particular abiotic factors influenced the grouping of sites.
Ad-hoc contrasts were used to identify differences between transplant groups, within a general linear model framework. Contrasts included (1) month dependent shifts in brightness between corals transplanted to and from the site of origin and (2) site and transplant dependent shifts in the mean brightness of a species. Seasonal patterns in mean brightness were identified with trigonometric linear modeling, applying a period of 12 to accommodate our monthly data interval.
3 Results
3.1 16-Year Comparison of Coral Communities
Analysis of the CREMP dataset revealed that coral cover declined at 8 of 10 reefs between 1996 and 2011 (Figure 2). Decreases in coral cover were greatest between 1999 and 2000. Coral cover declined from 25-15% at the three western-most inshore sites (Western Head, Cliff Green, and West Washerwoman) between 1999 and 2000 followed by a marginal rate of decline or stability in the remaining coral cover. The eastern most sites, Dustan Rocks and Western Turtle Shoal, maintained coral cover during this period while coral cover at all offshore bank reef sites decreased significantly between 1996 and 2000. Despite similar trends of coral cover loss, the total mean coral cover was significantly greater at the monitored inshore patch reefs compared to offshore bank reefs (RM-ANOVA: p < 0.0001).
.png) Figure 2 Annual percentage (%) of scleractinian coral cover from 1996-2011. Offshore shallow reef sites are grouped along the top row while inshore patch reef sites are grouped along the bottom row. The solid line is the mean across all years for inshore sites (14.6% coral cover) while the dotted line is the mean for offshore sites (6% coral cover). See figure 1 for site locations. |
In addition to the significant differences in coral cover, we identified community dissimilarities between offshore and inshore reefs. Species richness was significantly greater at the inshore sites (ANOVA: p < 0.05), with 9 additional species present when rare species were considered. Removing species accounting for less than 0.1% of the mean benthos cover resulted in a single species difference between inshore and offshore reefs. Rényi profiles of coral abundance indicated that all sites were dominated by a few species and therefore, are characterized by low species evenness (Figure 3). The most speciose sites, Looe Key from offshore reefs and Western Head from inshore reefs, displayed the least evenness. Offshore sites other than Looe Key reef, displayed similar profiles, indicated by the similar slopes of Rényi profiles. Although inshore sites displayed similar species richness, diversity profiles of sites within this zone indicate within zone community variation. Dustan Rocks and Western Washerwoman were the most similar of the inshore sites while Cliff Green with very similar richness to these two sites, was less even.
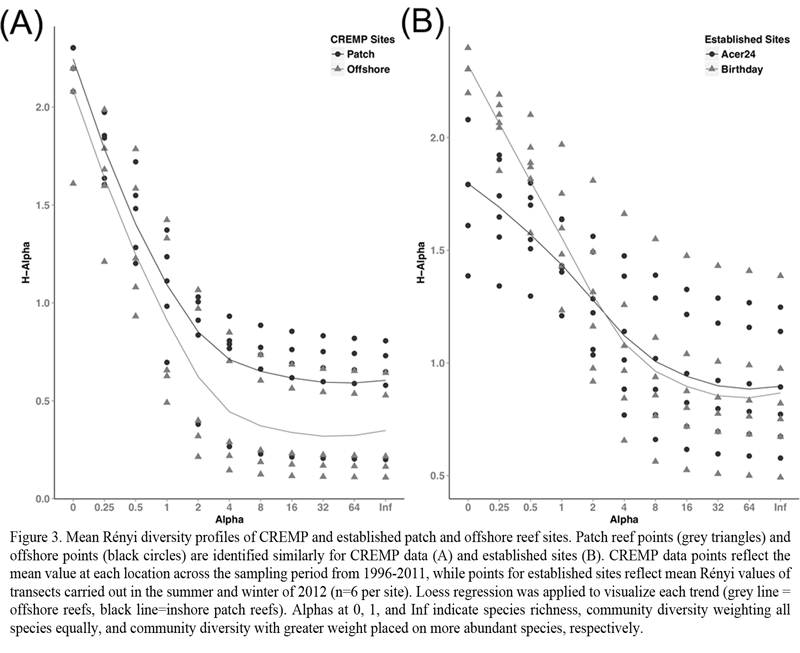 Figure 3 Mean Rényi diversity profiles of CREMP and established patch and offshore reef sites. Patch reef points (grey triangles) and offshore points (black circles) are identified similarly for CREMP data (A) and established sites (B). CREMP data points reflect the mean value at each location across the sampling period from 1996-2011, while points for established sites reflect mean Rényi values of transects carried out in the summer and winter of 2012 (n=6 per site). Loess regression was applied to visualize each trend (grey line = offshore reefs, black line=inshore patch reefs). Alphas at 0, 1, and Inf indicate species richness, community diversity weighting all species equally, and community diversity with greater weight placed on more abundant species, respectively. |
Zonal ordination of coral communities was identified following PCoA (Figure 4). Inshore sites ordinated closely together in quadrant 3 while offshore reefs, although displaying more variability, ordinated in the periphery away from this cluster in quadrants 1, 2, and 4, centering on quadrant 2 (Figure 4). Evaluating the loadings, along the first two axes, the abundances of some coral species were dissimilar between locations. Inshore patch reefs were characterized by an increased prevalence of M. cavernosa and Sidereastrea siderea, while P. astreoides and Acropora palamata were more common at offshore bank reefs.
Analysis of CREMP data supports the ongoing observation of increased abundance of corals at inshore patch reefs, however the CREMP data set is limited to relative abundance. To circumvent this issue we selected two reference sites within each zone to analyze in greater detail, focusing on coral community characteristics, particularly colony abundance, colony size, and benthic community composition. The mean seasonal benthos composition at Acer 24 reef (offshore) and Birthday reef (inshore) along 30 m transects is presented in Figure 5. Scleractinian coral cover was significantly greater at Birthday Reef (ANOVA: p < 3.83e-07), while the benthic community at Acer 24 contained significantly greater gorgonian coral cover (ANOVA: p < 2.01e-08), supporting our observations from the inshore vs. offshore analyses. Gorgonian cover also displayed seasonality at Acer 24 reef, with a greater percent cover observed during winter sampling (ANOVA: p < 0.003). This pattern was not evident at Birthday reef. Macro algal and turf algal cover was not significantly different between the Birthday reef and Acer 24, and accounted for greater than 30% of all reef cover. Increased macro algal cover was observed during winter months, but this difference was not significant compared to summer.
.png) Figure 4 Principal coordinate analysis of species occurrence from 1996-2011. The scores for each site (A) and loadings for each species (B) following principal coordinate analysis are presented. The first two coordinates accounting for 64.5% of variation in the data are plotted. Error bars (A) indicate 95% confidence intervals across the 15-year period. Species present in a particular quadrant of the loadings plot are more likely to be associated with a site in the same quadrant of the score plot. |
Coral community composition (Figure 3), species’ abundance (Figure 6), and colony size (Figure 7) were important components of the dissimilarity between the two sites. Rényi diversity profiles support the dissimilarity in scleractinian coral community composition (Figure 3). Specifically, species richness was greater at Birthday reef as indicated by an increased diversity value at alpha = 0. Fourteen scleractinian coral species were present at Birthday reef while only ten of these species were observed at Acer 24 reef. Additionally, both sites were characterized by low species evenness indicated by diversity values > 1 for alpha = Infinity. The most frequently observed colonies inhabiting Acer 24 reef were S. siderea, M. cavernosa, P. astreoides, and Stephananocoenia michelinii (Figure 6). In addition to these four species, Orbicella annularis was frequently observed at Birthday reef. Larger colonies were more frequently observed at Birthday Reef, as indicated by the greater positive skew of total coral frequency distributions (Figure 7). Frequency distributions were dependent upon zone for the three most common species, P. astreoides, S. siderea, and M. cavernosa.
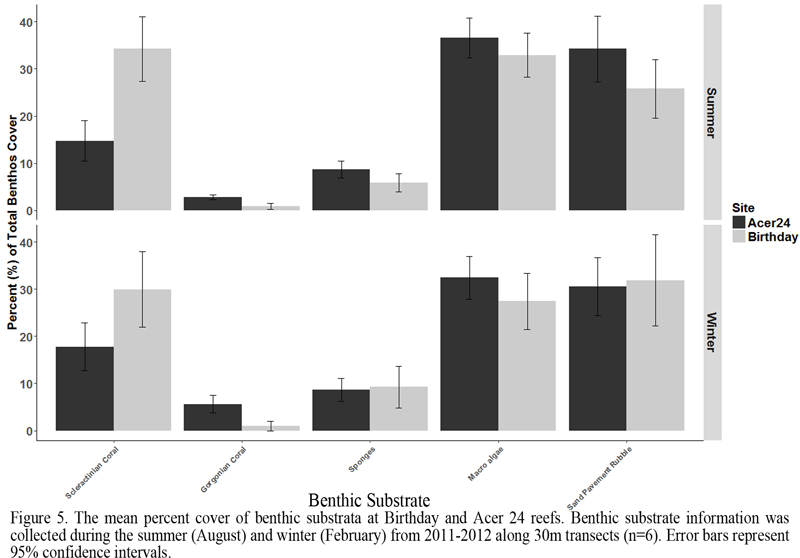 Figure 5 The mean percent cover of benthic substrata at Birthday and Acer 24 reefs. Benthic substrate information was collected during the summer (August) and winter (February) from 2011-2012 along 30m transects (n=6). Error bars represent 95% confidence intervals. |
3.2 Abiotic Characteristics
Principal component analysis (PCA) was applied to winter and summer subsets of the WQMP dataset to identify factors that may be associated with stress during these periods and influence the coral communities of inshore and offshore reefs. During winter months (January and February) 55% of the variation between sites was accounted for by three components (Figure 8). Variation along principal component 1 (22.9%) was influenced by water chemistry attributes. Total organic nitrogen (TON) ordinated sites negatively while the inorganic nutrients ammonium and silicate ordinated sites positively along this axis. These three variables did not vary significantly by region or location, however the marginal assurance in variation associated with silicate (ANOVA; p = 0.0652) provides a potential difference between inshore and offshore reefs during winter months. Along principal component 2 (19.2%), variation between sites was dictated by nitrate, soluble reactive phosphorus (SRP), chlorophyll a (Chl a), and total organic carbon (TOC). Nitrate was significantly greater at eastern sites than western sites and independent of location (ANOVA; p = 0.0062), while SRP, Chl a, and TOC remained statistically similar across regions and locations. The variation along principal component 3 (13.1%) was dictated by the variation in the physical characteristics turbidity and temperature. Turbidity was significantly greater at western sites regardless of location (ANOVA; p = 0.0113), while SWT was not significantly different for either region or location. Additionally, during winter months, greater variation in SWT was observed at inshore reefs (18-29ºC) than on offshore reefs (20-25ºC).
Plotting each of these components against one another resulted in regional ordination (Figure 8). With the exception of Rock Key, the western most sites West Washerwoman, Eastern Sambo, Cliff Green, Western Head and Sand Key, oriented away from eastern sites: Looe Key, Sombrero Reef, Dustan Rocks and Western Turtle Shoal. Variation in SWT, turbidity, and TOC appear responsible for the observed region dependent ordination during winter months, indicated by the direction and magnitude of arrows for each environmental variable. Within this structure, the three most speciose inshore sites, Western Head, Cliff Green, and West Washerwoman ordinated near one another.
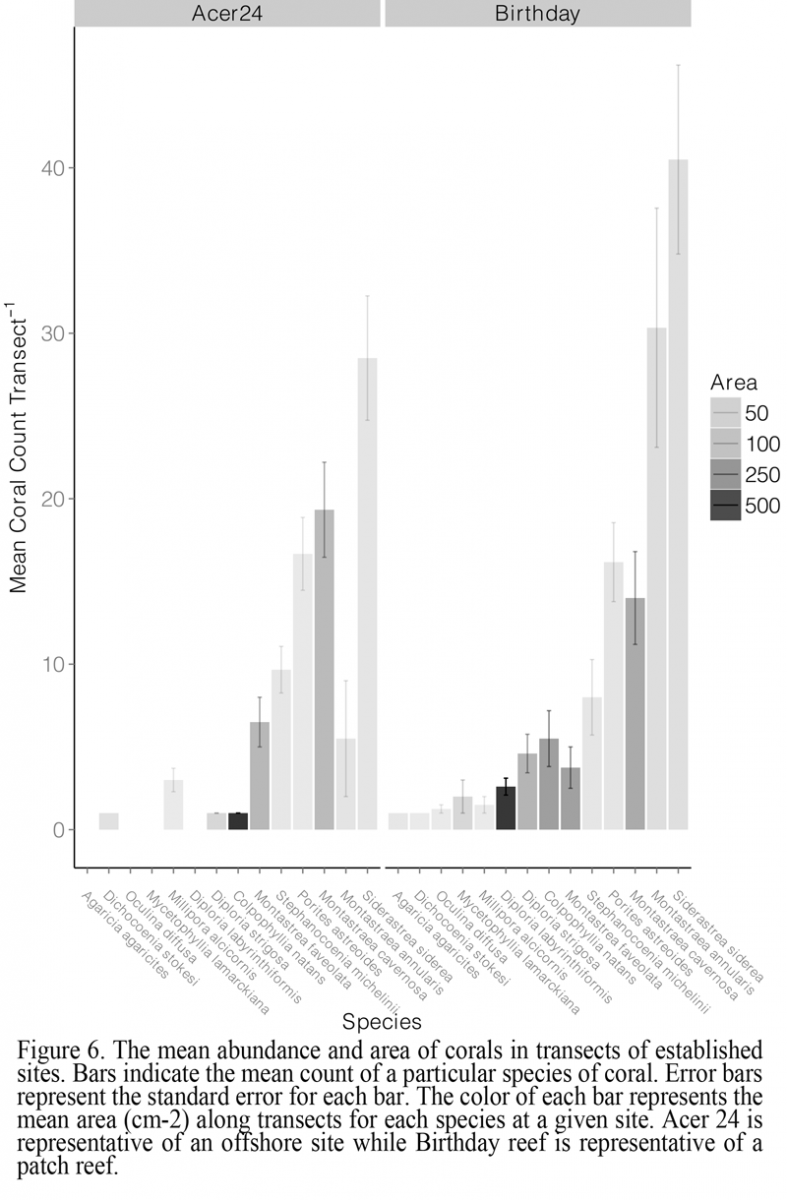 Figure 6 The mean abundance and area of corals in transects of established sites. Bars indicate the mean count of a particular species of coral. Error bars represent the standard error for each bar. The color of each bar represents the mean area (cm-2) along transects for each species at a given site. Acer 24 is representative of an offshore site while Birthday reef is representative of a patch reef. |
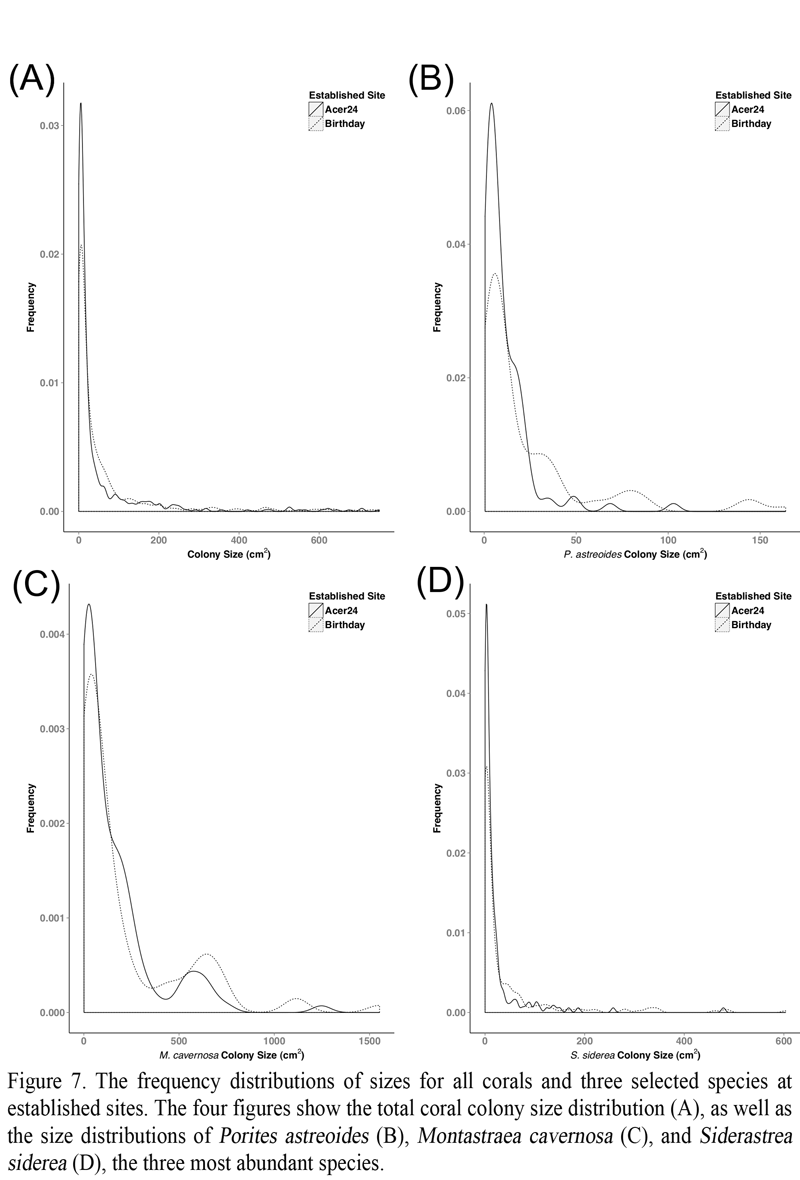 Figure 7 The frequency distributions of sizes for all corals and three selected species at established sites. The four figures show the total coral colony size distribution (A), as well as the size distributions of Porites astreoides (B), Montastraea cavernosa (C), and Siderastrea siderea (D), the three most abundant species. |
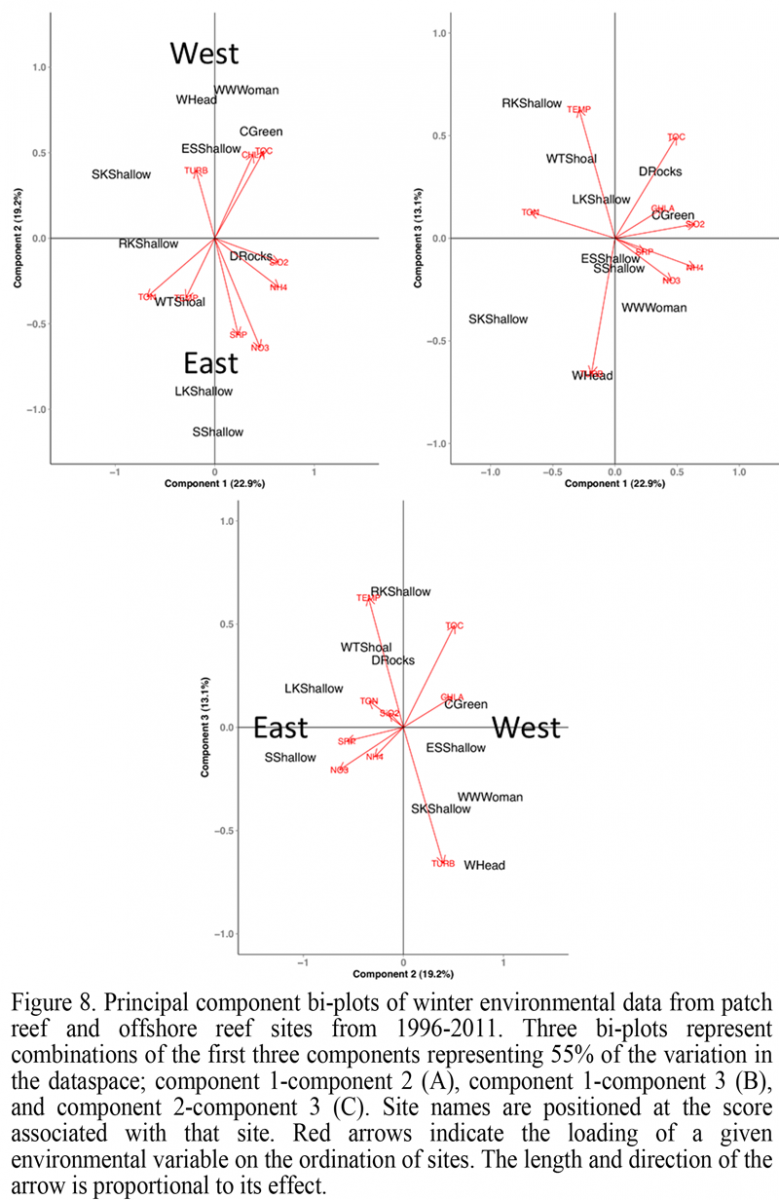 Figure 8 Principal component bi-plots of winter environmental data from patch reef and offshore reef sites from 1996-2011. Three bi-plots represent combinations of the first three components representing 55% of the variation in the dataspace; component 1-component 2 (A), component 1-component 3 (B), and component 2-component 3 (C). Site names are positioned at the score associated with that site. Red arrows indicate the loading of a given environmental variable on the ordination of sites. The length and direction of the arrow is proportional to its effect. |
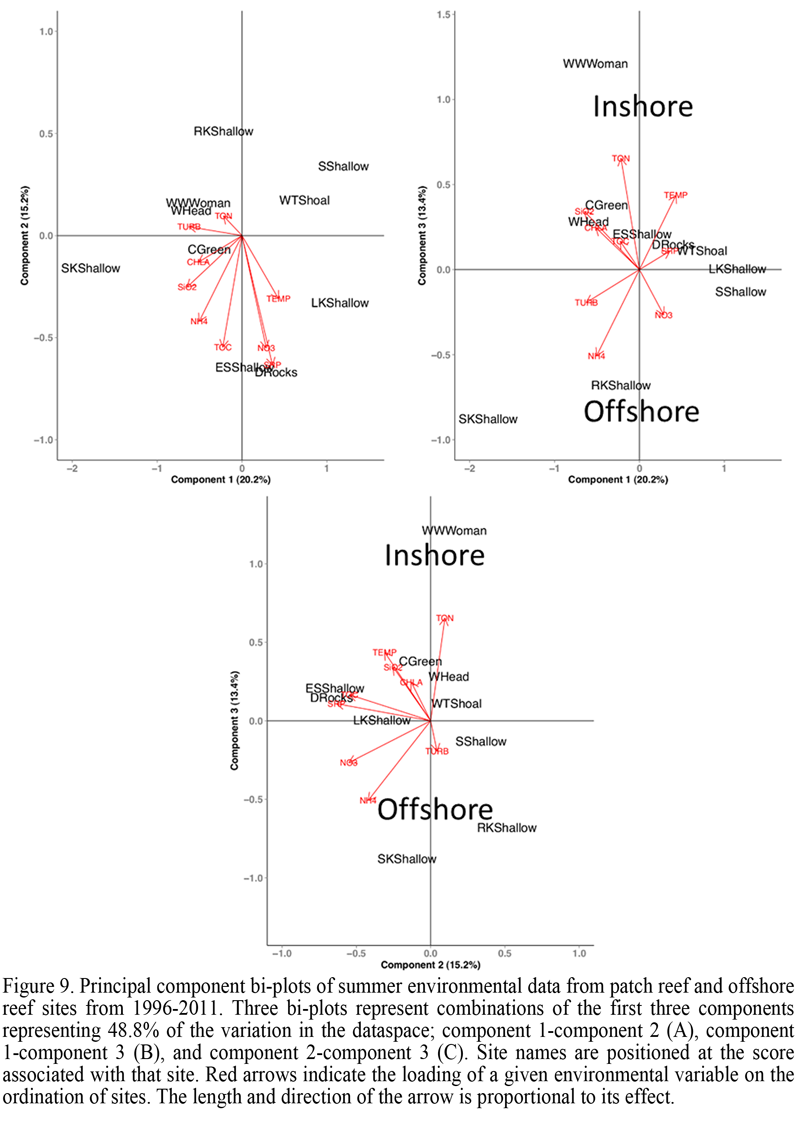 Figure 9 Principal component bi-plots of summer environmental data from patch reef and offshore reef sites from 1996-2011. Three bi-plots represent combinations of the first three components representing 48.8% of the variation in the dataspace; component 1-component 2 (A), component 1-component 3 (B), and component 2-component 3 (C). Site names are positioned at the score associated with that site. Red arrows indicate the loading of a given environmental variable on the ordination of sites. The length and direction of the arrow is proportional to its effect. |
Principal component analysis of summer WQMP data indicated 48.8% of the environmental variability among all sites could be explained by the first three components (Figure 9). Similar to PCA of winter months, regional ordination of sites was evident during the summer (Figure 9). The three speciose western sites Western Washerwoman, Cliff Green, and Western Head ordinated away from the eastern sites Sand Key, Rock Key and Sombrero Reef, and Looe Key, while the remaining sites, Eastern Sambo, Western Turtle Shoal, and Dustan Rocks were found between these two groups. This separation was not as distinct as that identified during winter months and may provide an indication of the large potential for site or location dependent environmental variability during summer months for reefs of the middle and lower Florida Keys.
Despite explaining only 13.3% of the total variation, PC3 appeared largely responsible for the regional ordination of sites within the data-space (Figure 9). Clear ordination was evident only in bi-plots including this component as an axis (Figure 9A, B). Therefore the environmental variables TON, ammonium, and SWT, which were most influential along PC3, may be important factors governing summer environmental differences between these regions and sites. Upon closer inspection, SWTs did not vary significantly between regions, however inshore sites were significantly warmer than offshore sites by 1°C (ANOVA; p = 0.0183). Further, SWT at inshore sites was consistently warmer during summer months (32-27°C), while offshore site SWTs were typically cooler but also ranged more widely than inshore sites during this time (31-20°C). Significant differences were not identified at the location or region level for TON (ANOVA; p = 0.166 and p = 0.214 respectively) or ammonium (ANOVA; p = 0.42 and p = 0.06 respectively).
Principal components 1 and 2, despite limited influence on the regional ordination of sites, imparted greater percentages of variation to the summer environment data-space than PC3. Influential variables along principal component 1 (20.2%) included factors that contribute to turbidity including Chl a and silicates. Western sites were 0.7 nephelometric turbidity units (NTU) more turbid than eastern sites (ANOVA; p = 0.022) and also significantly greater in Chl a (ANOVA; p = 0.013). Silicates displayed significant variation at the zone (ANOVA; p = 0.0392) but not regional level. Variation along PC2 was influenced by SRP, nitrate, and TOC, however significant variation was not identified for any of these variables at the regional or site level.
Increasing the resolution of temperature data to daily monitoring at a reference inshore and offshore site provided greater support to these findings. During three years of temperature monitoring (2011-2013) annual mean SWT at the inshore site, Birthday reef, and offshore site, Acer 24 reef, did not significantly differ (ANOVA p = 0.828); 26.74°C and 26.77°C respectively. Monthly variance in SWT, however was 0.5°C less at Acer 24 (ANOVA p < 0.05). Additionally increased (> 30°C) and decreased (< 23°C) daily SWTs occurred more frequently (55%) at Birthday reef compared to Acer 24.
3.3 Location-Dependent Shifts in Brightness, a Bleaching Related Characteristic
Fragments of M. cavernosa and P. astreoides transplanted to Birthday reef displayed significantly fewer signs of bleaching (i.e. increased brightness) at Birthday reef (ANOVA; p = 2e-16), compared to the conspecifics transplanted to the offshore site, Acer 24 (Figure 10). Although the occurrence of severely bleached corals (Brightness ≥ 150) was rare, the frequency of a lesser degree of bleaching (101 ≤ Brightness ≥ 149) was significantly greater at the offshore site, Acer 24. We did not observe a transplant dependent effect on the brightness of corals when analyzing the total frequency of occurrences at the three brightness ranges. Observing the quantum yield of photochemical energy conversion (Yield: ΔF/Fmˊ) of corals inhabiting each location, we found significant differences at the species (p = 2.23e-06) and site (p = 0.0198) levels during summer months (Figure 11). During the summer the probability of photons entering photosystem II (PSII) was greater for P. astreoides and colonies of these two species inhabiting Birthday reef. Corals inhabiting Birthday reef did not display differences in yields between winter and summer.
We observed a seasonal change in the monthly brightness of coral fragments transplanted to Birthday reef, the inshore site, following the application of a Loess smoothing function (Cleveland and Devlin, 1988). The lightest shade (greater brightness value) was observed during September and October while the darkest (lower brightness value) were observed during February and March. Due to the large variance in monthly brightness values between coral fragments, trigonometric linear regression was applied to determine if a significant trend existed. Changes in brightness significantly fit a cosine function with 12-month period (ANOVA; p = 0.04), representative of a significant annual pattern in coral brightness at Birthday reef (Figure 12). A significant cosine pattern was not observed for corals at Acer 24. Instead, linear regression of coral fragment brightness at this site indicated that mean brightness values of corals transplanted to this site increased from September 2011 to May 2013. Therefore these corals became progressively lighter (potentially more bleached) during this period of time.
4 Discussion
The inshore patch reef system of the FKNMS has maintained stable and productive scleractinian coral communities since the last large scale decreases in coral cover around 1999. The neighboring bank reef communities experienced even more dramatic losses to coral cover prior to 1999 and have yet to rebound (Lirman and Fong, 2007; Schutte et al., 2010). Our analysis of the past 16 years of data from CREMP indicates that inshore reefs continue to support the majority of coral cover in the Lower Florida Keys (Figure 2). Analysis of the 10 reefs from the CREMP indicated that the inshore patch reef system now accounts for greater than 70% of the coral cover remaining throughout the FKNMS regardless of the more consistent environment associated with offshore reefs. We observed a greater percentage of live coral cover and greater coral diversity at five sites within the inshore reef system spanning the lower and middle keys compared to five bank reef sites. Community level analysis of an inshore and offshore reef site, further support that scleractinian corals inhabiting inshore sites are not only more abundant (Figure 2) and more diverse (Figure 3), but mean colony size is significantly greater (Figure 7). Although the rapid loss of dominant acroporid corals from the bank reef system caused a dramatic shift in reef biota (Patterson et al., 2002) as a consequence of the epizootic pathogen Serratia marsescens (Patterson et al., 2002; Sutherland et al., 2011), this event alone does not explain the failure of the currently dominant coral species (Burman et al., 2012) to re-establish this zone over the past 20 years given the apparent absence of other potential environmental constraints.
.png) Figure 10 Frequency of occurrence for three brightness states following a reciprocal transplant of P. astreoides and M. cavernosa. Three bins were created for mean brightness values, each spanning approximately 40 units. The four graphs are separated along the y-axis by the site to which a coral was transplanted. Bar color indicates collection site (Black = Acer 24, Grey = Birthday). The graphs are further separated by species along the x-axis. Bar height is indicative of the cumulative number of occurrences within a particular bin from monthly images taken from September 2011-April 2013. |
Our analysis indicates that inshore and offshore reefs of similar depth (6 m) within the middle and lower FKNMS support distinct scleractinian coral communities. The abundances of four species greatly influenced the ordination of inshore sites away from offshore sites; Acropora palmata, S. siderea, M. cavernosa, and P. astreoides (Figure 4). The influence of A. palmata was least revelatory as this known specialist inhabits a narrow habitat range including high relief and inner line spur and groove habitats found only in the offshore reef zone (Miller et al., 2008). The current endangered status of A. palmata also precluded it’s inclusion in our transplant study. Siderastrea siderea, the most common scleractinian coral currently inhabiting the FKNMS (Burman et al., 2012), was highly prevalent regardless of the zone. Because the goal of this study was to identify sources of variation in the coral cover of inshore and offshore sites, the known generalist nature of S. siderea (Burman et al., 2012) precluded this species as well. The remaining two species, P. astreoides and M. cavernosa, although considered eurytopic (Burman et al., 2012), inhabited inshore and offshore zones with differing degrees of prevalence suggesting dissimilar tolerances to the stressors inherent to each zone. We selected these species as a proxy to identify site and species dependent factors that may contribute to the continued trend of decreased coral cover at offshore compared to inshore reefs.
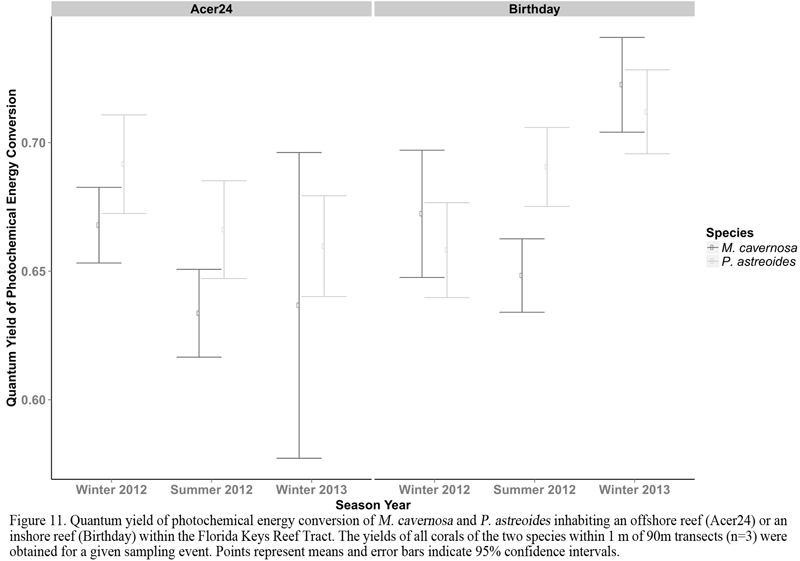 Figure 11 Quantum yield of photochemical energy conversion of M. cavernosa and P. astreoides inhabiting an offshore reef (Acer24) or an inshore reef (Birthday) within the Florida Keys Reef Tract. The yields of all corals of the two species within 1 m of 90m transects (n=3) were obtained for a given sampling event. Points represent means and error bars indicate 95% confidence intervals. |
Reciprocal transplantation of M. cavernosa and P. astreoides between Birthday Reef (inshore site) and Acer 24 (offshore site), revealed that fragments of both species transplanted to the inshore site, displayed significantly lower brightness in images than conspecifics and ramets transplanted to the offshore site, regardless of their collection site (Figure 11). We interpret this result to signify decreased levels of stress or photoacclimation at inshore reefs resulting in greater symbiont concentrations within the host or increased pigment concentration within the algal symbionts (Symbiodinium spp. commonly referred to as zooxanthellae). Although increases in symbiont density can also occur when inorganic nitrogen concentrations increase (Muscatine et al., 1989; Marubini and Davies, 1996), our study of the abiotic differences between the inshore and offshore zone did not yield a significant difference in inorganic nitrogen forms between zones. A sister study examining the symbiont diversity within the corals of this study determined that symbiont diversity was greater for corals collected from the offshore reef but this diversity was not affected by where a fragment was transplanted to (Hauff et al., 2016). Instead, inshore sites displayed increased seawater temperatures and a wider range of turbidities. Further, the two CREMP sites nearest to our transplant sites, Western Washerwoman (inshore) and Looe Key (offshore), displayed significantly different turbidities during the winter (µ = 3.1 NTU and µ = 0.26 NTU respectively) and summer following analysis of the WQMP dataset (µ = 0.8218 NTU and µ = 0.480 NTU respectively). From this information differences in brightness of transplanted coral colonies may be a function of the effects of turbidity and temperature. Temperature and turbidity have been identified as mediators of growth for Orbicella annularis populations near Key Largo, FL (Hudson, 1981).
While transplantation site affected the brightness of coral fragments, the conditions previously experienced by a transplanted fragment (i.e. collection site) did not have a significant effect on this observation. It is important to caution that changes in brightness may be caused by an array of factor in corals in including host pigments, symbiont loss, symbiont pigments, and the environment. While we controlled for site dependent differences as best as possible these factors could contribute to the observed variability. Evidence that the differences in abiotic conditions identified between these two sites were not significant enough to elicit a visible response following transplantation. This finding is contradictory to studies that have reported significant interactions between the environmental history experienced by these two species and their response to future stress (Haslun et al., 2011; Kenkel et al., 2013). For instance, we have shown previously (Haslun et al., 2011) that M. cavernosa from the Flower Garden Banks National Marine Sanctuary (FGBNMS), a well-developed and thermally stable scleractinian reef ecosystem, are more susceptible to increased thermal stress than conspecifics from the FKNMS, where increased SWTs are experienced. Similarly, P. astreoides has been shown to display experience-dependent acclamatory responses via the expression of genes associated with calcification and metabolism when exposed to different temperature regimes (Kenkel et al., 2013). Both of the aforementioned studies reported that pronounced differences in seawater temperature existed between the collection locations. In this study, mean maximum and mean minimum temperatures for summer and winter respectively, deviated by approximately 1°C between inshore and offshore zones, which is less than that observed between the FGBNMS and the FKNMS (Haslun et al., 2011) or the more extreme temperature regimes at near shore sites (< 1 km from shore) of the FKNMS (Kenkel et al., 2013). It is possible that the dissimilarity in SWT observed between our sites was not acute enough to illicit an acclamatory effect in the observed brightness of P. astreoides and M. cavernosa or another stressor that was unmeasured imparted the perceived responses.
Bleaching (loss of color largely associated with symbionts) of corals during summer seasons is a common response to increased stress from light and temperature levels (Fitt et al., 2000) and not necessarily detrimental to the organism. The primary purpose of this response is to limit the host’s cellular stress caused by symbiont malfunction (Strychar et al., 2004). Coral fragments experiencing the conditions at the inshore site displayed seasonality in colony brightness, characterized by a summer stress period and a winter compensatory period (Figure 12). The seasonal pattern was also reflected in the significant relationship identified between temperature and brightness for these corals. Therefore, seasonal stress levels from temperature remain coupled to bleaching at Birthday reef and may be critical to the continued success of coral at inshore reefs via local adaptation or acclimatization to the conditions present. Seasonality in coral fragment brightness was not detected for fragments transplanted to Acer 24. On the contrary, we observed a significant positive linear trend between brightness and month, indicating that fragments of both species of coral became progressively lighter over this two-year period. Therefore, bleaching (the loss of symbionts) and temperature stress can be considered uncoupled for P. astreoides and M. cavernosa experiencing the conditions at Acer 24. Given the narrower annual temperature range characteristic of this site, this result is intriguing. From our results we hypothesize that stress resulting from increased irradiance (i.e. decreased turbidity), characteristic of offshore sites, provides an additional source of stress increasing the cumulative stress level during summer and winter periods beyond that of corals at inshore sites. The increased cumulative stress level may decrease a colony’s potential for recovery during the winter season resulting in a state of chronically mild bleaching (i.e. more pale than conspecifics).
Chronic bleaching (an archetype of chronic distress) can be difficult to observe given its non-lethal nature (Lasker and Coffroth, 1999). Horizontal linear extension was evident at both transplant sites throughout the course of our study (pers. obs.), however we were unable to determine whether or not the degree of growth differed between sites. In order to detect potential differences in fitness associated with chronic distress, we investigated site dependent community characteristics. Eventual mortality may occur when organisms must allocate resources to protection and maintenance (Lasker and Coffroth, 1999) rather than fecundity and growth. We observed a significant difference between the mean colony area of M. cavernosa and P. astreoides at Birthday reef and Acer 24, as well as another commonly observed species, S. siderea (a known generalist). Similar observations have been noted outside of the Florida Keys (Edmunds and Elahi, 2007). Following dramatic decreases in the abundance of O. annularis (formerly Montastraea annularis) at a reef near St. John Island in the United States Virgin Islands, the mean size of corals continued to decrease over a five-year period, from 1999-2003, despite the stabilization of percent coral cover (Edmunds and Elahi, 2007). The authors projected that this species of coral would become locally extinct if the level of stressors remained unchanged over the next 30-50 years. Corals at offshore reefs of the FKNMS appear to be experiencing this trend now.
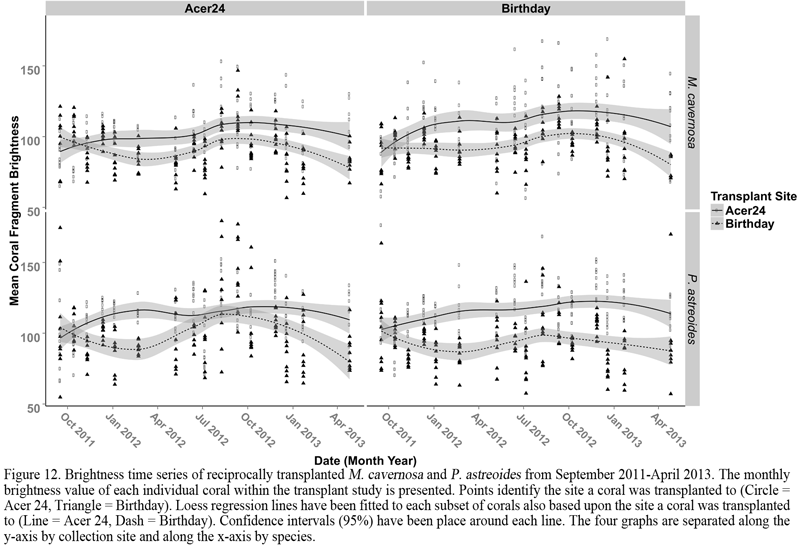 Figure 12 Brightness time series of reciprocally transplanted M. cavernosa and P. astreoides from September 2011-April 2013. The monthly brightness value of each individual coral within the transplant study is presented. Points identify the site a coral was transplanted to (Circle = Acer 24, Triangle = Birthday). Loess regression lines have been fitted to each subset of corals also based upon the site a coral was transplanted to (Line = Acer 24, Dash = Birthday). Confidence intervals (95%) have been place around each line. The four graphs are separated along the y-axis by collection site and along the x-axis by species. |
5 Conclusion
Offshore bank reefs harbor relict communities of scleractinian corals and these communities do not show signs of rebounding. Although the stressors associated with this zone appear lower than those associated with more successful inshore patch reefs our results indicate that corals inhabiting offshore shallow reefs are experiencing more significant levels of stress compared to conspecifics from inshore patch reefs. This result may not be reflected in the acuteness of stress but rather the absence of a phase to recover from seasonal stressors. In support of this claim we provide several tracts of evidence. First, the brightness of corals (an indication of symbiont density) transplanted to an offshore site became progressively lighter over a two-year period regardless of the site of collection while corals transplanted to an inshore site displayed seasonality in brightness. Second, the linear relationship between seawater temperature and brightness was significant for corals transplanted to inshore sites but this relationship did not exist for corals transplanted to an offshore site. Third, the mean colony size of the two species used for transplantation as well as a common generalist, were smaller at offshore sites. Finally, we identified a factor, turbidity that provides an explanation supporting the potential for chronic stress to occur at offshore sites. We propose that decreased turbidity results in increased irradiance at offshore sites resulting in greater cumulative stress than is present at inshore reefs. The results presented in this study provide support for increased protection and awareness of inshore patch reefs. It also provides important information regarding the important synergism between changing temperature and irradiance experienced by corals. Increased turbidity provides both a refuge from increased irradiance as well as a potential food source to counteract decreases in algal carbon fixation for the cnidarian host during periods of eutrophication as long as sedimentation is not significant (Bongiorni et al., 2003; Cacciapaglia and van Woesik, 2016).
Author Contributions
JAH designed and conducted the experiments and statistical analysis. BH designed and conducted the experiments. KBS designed and advised experiments, and provided laboratory space for the work. JMC advised work and provided space for samples. All authors read and approved the final manuscript.
Acknowledgments
First and foremost we would like to thank the staff of the Mote Marine Laboratory Tropical Research Station for their help in throughout the course of this research. We would also like to thank the Conservation Protection Network for their continued dedication to and support of this research. Finally thanks to the Annis Water Resources Institute for their support.
Alvarez-Filip L., Dulvy N.K., Gill J.A., Côte I.M., and Watkinson A.R., 2009, Flattening of Caribbean coral reefs: region-wide declines in architectural complexity. Proceedings of the Royal Society Biology. 276: 3019-3025.
http://dx.doi.org/10.1098/rspb.2009.0339
Aronson R.B., and Precht W.F., 2001, White-band disease and the changing face of the Caribbean coral reefs. Hydrobiologia 460: 25-38.
http://dx.doi.org/10.1023/A:1013103928980
Bongiorni L., Shafir S., and Rinkevich B., 2003, Effects of particulate matter released by a fish farm (Eilat, Red Sea) on survival and growth of Stylophora pistillata coral nubbins. Marine Pollution Bulletin. 46: 1120-1124.
http://dx.doi.org/10.1016/S0025-326X(03)00240-6
Buddemeier R.W., and Fautin D.G., 1993, Coral bleaching as an adaptive mechanism. Bioscience. 43: 320-326.
http://dx.doi.org/10.2307/1312064
Burman S.G., Aronson R.B., and van Woesik R., 2012, Biotic homogenization of coral assemblages along the Florida reef tract. Marine Ecological Progress Series. 467: 89-96.
http://dx.doi.org/10.3354/meps09950
Cacciapaglia, C., van Woesik, R., 2016, Climate-change refugia: shading reef corals by turbidity. Global Change Biology. 22(3): 1145:1154.
Cervino J.M., Hauff B., Haslun J.A., Winiarski-Cervino K., and Cavazos M., 2012, Ulcerated yellow spot syndrome: implications of aquaculture-related pathogens associated with soft coral Sarcophyton ehrenbergi tissue lesions. Diseases of Aquatic Organisms. 102: 137-148.
http://dx.doi.org/10.3354/dao02541
Cleveland W.S., and Devlin S.J., 1988, Locally-weighted fitting: an approach to fitting analysis by local fitting. Journal of American Statistical Association. 83: 596-610.
http://dx.doi.org/10.1080/01621459.1988.10478639
Computing. R Foundation for Statistical Computing, Vienna, Austria. 2013;
Coyer J.A., Steller L.D., and Witman J, The underwater catalog: a guide to methods in underwater research. 2nd ed. Ithaca, New York: Shoals Marine Laboratory; 1999.
Edmunds P.J., and Elahi R., 2007, The demographics of a 15-year decline in cover of the Caribbean reef coral Montastraea annularis. Ecological Monographs. 77(1): 3-18.
http://dx.doi.org/10.1890/05-1081
Fitt W.K., McFarland F.K., Warner M.E., and Chilcoat G.C., 2000, Seasonal patterns of tissue biomass and densities of symbiotic dinoflagellates in reef corals and relation to coral bleaching. Limnology and Oceanography. 45(3): 677-685.
http://dx.doi.org/10.4319/lo.2000.45.3.0677
Foster N.L., Paris C.B., Kool J.T., Baums I.B., Stevens J.R., et al., 2012, Connectivity of Caribbean coral populations: complementary insights from empirical and modelled gene flow. 21: 1143-1157.
Genty B., Briantais J.M., and Baker N.R., 1989, The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence; Biochimica Biophysica Acta. 990: 87-92.
http://dx.doi.org/10.1016/S0304-4165(89)80016-9
Ginsburg R.N., Gischler E., and Kiene W.E., 2001, Partial mortality of massive reef-building corals: An index of patch reef condition, Florida reef tract. Bulletin of Marine Science. 69(3): 1149-1173.
Guest J.R., Baird A.H., Maynard J.A., Muttaqin E., and Edwards A.J., 2010, Contrasting patterns of coral bleaching susceptibility in 2010 suggest an adaptive response to thermal stress. PLOS ONE. doi:10.1371/journal.pone.0033353.
http://dx.doi.org/10.1371/journal.pone.0033353
Haslun J.A., Strychar K.B., Buck G., and Sammarco P.W., 2011, Coral bleaching susceptibility is decreased following short-term (1-3 year) prior temperature exposure and evolutionary history. Journal of Marine Biology. Article ID 406812: 1-13.
Hsu C., de Palmas S., Kuo C., Denis V., and Chen C.A., 2014, Identification of scleractinian coral recruits using fluorescent censusing and DNA barcoding techniques. PLOS ONE. 9(9): 1-13.
http://dx.doi.org/10.1371/journal.pone.0107366
Hudson J.H., 1981, Growth rates in Montastraea annularis: a record of environmental change in Key Largo Coral Reef Marine Sanctuary, Florida. Bulletin of Marine Science. 31(2): 444-459.
Hudson J.H., Shinn E.A., Halley R.B., and Lidz B., 1976, Slerochronology: a tool for interpreting past environments. Geology. 4: 361-364.
http://dx.doi.org/10.1130/0091-7613(1976)4%3C361:SATFIP%3E2.0.CO;2
Jackson, J.E., 1980, Principal components and factor analysis: part I-principal components. Journal of Quality Technology. 12(4):201-213.
Jost L., 2006, Entropy and diversity. Oikos. 113(2): 363-375.
http://dx.doi.org/10.1111/j.2006.0030-1299.14714.x
Kenkel C.D., Meyer E., and Matz M.V., 2013, Gene expression under chronic heat stress in populations of the mustard hill coral (Porites astreoides) from different thermal environments. Molecular Ecology. 22(16): 4322-4334.
http://dx.doi.org/10.1111/mec.12390
Kohler K.E., and Gill S.M., 2006, Coral point count with excel extensions (CPCe): a visual basic program for the determination of coral and substrate coverage using random point count methodology. Computational Geoscience. 32: 1259-1269.
http://dx.doi.org/10.1016/j.cageo.2005.11.009
Lapointe B.E., Barile P.J., and Matzie W.R., 2004, Anthropogenic nutrient enrichment of seagrass and coral reef communities in the Lower Florida Keys: discrimination of local versus regional nitrogen sources. Journal of Experimental Marine Biology and Ecology. 308: 23-58.
Lasker H.R., and Coffroth M.A., 1999, Responses of clonal reef taxa to environmental change. American Zoologist. 39: 92-103.
http://dx.doi.org/10.1093/icb/39.1.92
Leujak W., and Ormond R.F.G., 2007, Comparative accuracy and efficiency of six coral community survey methods. Journal of Experimental Marine Biology and Ecology. 351(1-2): 168-187.
http://dx.doi.org/10.1016/j.jembe.2007.06.028
Lidz B.H., Reich C.D., and Shinn E.A., 2007, Systematic mapping of bedrock and habitats along the Florida Reef Tract – Central Key Largo to Halfmoon Shoal (Gulf of Mexico). U.S. Geological Survey Professional Paper. 1751: http://pubs.usgs.gov/pp/2007/1751/index.html.
Lirman D., and Fong P., 2007, Is proximity to land-based sources of coral stressors an appropriate measure of risk to coral reefs? An example from the Florida Reef Tract. Marine Pollution Bulletin. 54(6): 779-791.
http://dx.doi.org/10.1016/j.marpolbul.2006.12.014
Maliao R.J., Turingan R.G., and Lin J., 2008, Phase-shift in coral reef communities in the Florida Keys National Marine Sanctuary (FKNMS), USA. Marine Biology. 154: 841-853.
http://dx.doi.org/10.1007/s00227-008-0977-0
Marubini F., and Davies P.S., 1996, Nitrate increases zooxanthellae population density and reduces skeletogenesis in corals. Marine Biology. 127: 319-328.
http://dx.doi.org/10.1007/BF00942117
Miller M.W., Weil E., and Szmant A.M., 2000, Coral recruitment and juvenile mortality as structuring factors for reef benthic communities in Biscayne National Park, USA. Coral Reefs. 19: 115-123.
http://dx.doi.org/10.1007/s003380000079
Miller S.L., Chiappone M., Rutten L.M., and Swanson D.W., 2008, Population status of Acropora corals in the Florida Keys. Proc 11th International Coral Reef Symposium. 1:775-779.
Minchin P.R., O'Hara R.B., et al., 2013, vegan: Community Ecology
Moulding A.L., 2005, Coral recruitment patterns in the Florida Keys. International Journal of Tropical Biology. 53: 75-82.
Mumby P.J., and Steneck R.S., 2008, Coral reef management and conservation in light of rapidly evolving ecological paradigms. Trends in Ecology and Evolution. 23(10): 555-563.
http://dx.doi.org/10.1016/j.tree.2008.06.011
Muscatine L., Falkowski P.G., Dubinsky Z., Cook P.A., McCloskey L.R., 1989, The effect of external nutrient resources on the population dynamics of zooxanthellae in a reef coral. Proceedings of the Royal Society London Biology. 236(1284): 311-324.
http://dx.doi.org/10.1098/rspb.1989.0025
Noreen A.M., Harrison P.L., and Van Oppen M.J.H., 2009, Genetic diversity and connectivity in a brooding reef coral at the limit of its distribution. Proceedings of the Royal Society Biology. 276: 3927-3935.
http://dx.doi.org/10.1098/rspb.2009.1050
Oksanen F., Blanchet G., Kindt R., Legendre P., Package. R package version 2.0-10. 2013; http://CRAN.R-project.org/package=vegan.
Palandro D.A., Andréfouët S., Hu C., Hallock P., Müller-Karger F.E., and Dustan P., 2008, Quantification of two decades of shallow-water coral reef habitat decline in the Florida Keys National Marine Sanctuary using Landsat data (1984-2002). Remote Sensing of the Environment. 8(112): 3388-3399.
http://dx.doi.org/10.1016/j.rse.2008.02.015
Palumbi S.R., Barshis D.J., Traylor-Knowles N., and Bay R.A., 2014, Mechanisms of reef coral resistance to future climate change. Science. 344(6186): 895-898.
http://dx.doi.org/10.1126/science.1251336
Pandolfi J.M., Jackson J.B.C., Baron N., Bradbury R.H., Guzman H.M., Hughes T.P., et al., 2005, Are U.S. coral reefs on the slippery slope to slime? Science. 307(5716): 1725-1726.
http://dx.doi.org/10.1126/science.1104258
Patterson K.L., Porter J.W., Ritchie K.B., Polson S.W., Mueller E., Peters E.C., et al., 2002, The etiology of white pox, a lethal disease of the Caribbean elkhorn coral, Acropora palmata. Proceedings of the National Academy of Science. 99: 8725-8730.
http://dx.doi.org/10.1073/pnas.092260099
R Core Team. R: A language and environment for statistical
Roberts H.H., Rouse L.J., Walker N.D., and Hudson J.H., 1982, Coldwater stress in Florida Bay and northern Bahamas: a product of winter cold-air outbreaks. Journal of Sediment Petroleum. 52: 145-155.
Ruzicka R.R., Colella M.A., Porter J.W., Morrison J.M., Kidney J.A., Brinkhuis V., et al., 2013, Temporal changes in benthic assemblages on Florida Keys reefs 11 years after the 1997/1998 El Nino. Marine Ecology Progress Series. 489: 125-141.
http://dx.doi.org/10.3354/meps10427
Sammarco P.W., and Andrews J.C., 1989, The helix experiment: differential localized dispersal and recruitment patterns in Great Barrier Reef corals. Limnology and Oceanography. 34(5): 896-912.
http://dx.doi.org/10.4319/lo.1989.34.5.0896
Schmidt-Roach S., Kunzmann A., and Arbizu P.M., 2008, In situ observation of coral recruitment using fluorescence census techniques. Journal of Experimental Marine Biology and Ecology. 367(1): 37-40.
http://dx.doi.org/10.1016/j.jembe.2008.08.012
Schutte V.G.W., Selig E.R., and Bruno J.F., 2010, Regional spatio-temporal trends in Caribbean coral reef benthic communities. Marine Ecology Progress Series. 402: 115-122.
http://dx.doi.org/10.3354/meps08438
Shearer T.L., and Coffroth M.A., 2006, Genetic identification of Caribbean scleractinian coral recruits at the Flower Garden Banks and the Florida Keys. Marine Ecological Progress Series. 306: 133-142.
http://dx.doi.org/10.3354/meps306133
Shinn E.A., Reich C.D., Hickey T.D., and Lidz B.H., 2003, Staghorn tempestites in the Florida Keys. Coral Reefs. 22: 91-97.
http://dx.doi.org/10.1007/s00338-003-0289-2
Siebeck U.E., Marshall N.J., Klüter A., and Hoegh-Guldberg O, 2006, Monitoring coral bleaching using a color reference card. Coral Reefs 25: 453-460. Springer, New York
http://dx.doi.org/10.1007/s00338-006-0123-8
Soto I.M., Muller-Karger F.E., Hallock P., and Hu C., 2011, Sea surface temperature variability in the Florida Keys and its relationship to coral cover. Journal of Marine Biology. doi:10.1155/2011/981723.
http://dx.doi.org/10.1155/2011/981723
Steneck R.S., Paris C.B., Arnold S.N., Ablan-Lagman M.C., Alcala A.C., Butler M.J., et al., 2009, Thinking and managing outside the box: coalescing connectivity networks to build region-wide resilience in coral reef ecosystems. Coral Reefs. 28: 367-378.
http://dx.doi.org/10.1007/s00338-009-0470-3
Strychar K.B., Coates M., Sammarco P.W., and Piva T.J., 2004, Bleaching as a pathogenic response in scleractinian corals, evidenced by high concentrations of apoptotic and necrotic zooxanthellae. Journal of Experimental Marine Biology and Ecology. 304(1): 99-121.
http://dx.doi.org/10.1016/j.jembe.2003.11.023
Sutherland K.P., Shaban S., Joyner J.L., Porter J.W., and Lipp E.K., 2011, Human pathogen shown to cause disease in the threatened elkhorn coral Acropora palmata. PLoS ONE. 6(8): e23468. doi:10.1371/journal.pone.0023468.
http://dx.doi.org/10.1371/journal.pone.0023468
Walker N.D., Roberts H.H., Rouse L.J., and Huh O.K., 1982, Thermal history of reef-associated environments during a record cold-air outbreak event. Coral Reefs. 1: 83-87.
http://dx.doi.org/10.1007/BF00301689
Wickham H. ggplot2: elegant graphics for data analysis. 2009.
Ziemann D.A., Walsh, W.A., Saphore E.G., Fulton-Bennett K., 1992, A survey of water quality characteristics of effluent from Hawaiian aquaculture facilities. Journal of the World of Aquaculture Society. 23(3): 180-191.



. PDF(1849KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Joshua Haslun
. Briana Hauff
. Kevin Strychar
. James Cervino
Related articles
. Acclimation
. Coral
. Florida Keys
. Patch Reef
. Symbiodinium
Tools
. Email to a friend
. Post a comment