Particular Qualities of Identification and Taxonomy of Some Scleractinian (Scleractinia: Faviina), Faviidae Gregory, 1900 
 Author
Author  Correspondence author
Correspondence author
International Journal of Marine Science, 2016, Vol. 6, No. 14 doi: 10.5376/ijms.2016.06.0014
Received: 29 Jan., 2016 Accepted: 27 Feb., 2016 Published: 25 Mar., 2016
Latypov, 2016, Particular Qualities of Identification and Taxonomy of Some Scleractinian Faviidae Gregory, 1900, (Scleractinia: Faviina), International Journal of Marine Science, 6(14):1-10 (doi: 10.5376/ijms.2016.06.0014)
It is a brief history of taxonomy and identification of one of the most interesting groups of massive hermatypic corals family Faviidae, characterized by placoid, cerioid and meandroid forms of corallum and having with Acroporidae and Fungiidae the greatest number of genera and species diversity among all hermatypic Scleractinian. Describes terminology and morphological signs under the definition and description of corals of the family.
1 Introduction and Brief History of Taxonomy
The Faviidae is among the most important families of hermatypic corals, having after Acroporidae, the most number of genera and species among all scleractinian. Due to their abundance in the majority of reef biotopes throughout the entire Indo-Pacific, they are present from the 18th century in the most of taxonomically important collections. In one of the earliest taxonomic publications (Forskål, 1775) 23 scleractinian coral species from the Red Sea were described, of which 8 species were attributed to Faviidae. The ubiquity and the great diversity of these corals on the Vietnamese reefs (including unknown new species for science, Latypov, 2012, 2013), consider particularities of their identification and taxonomy. The taxonomic history of this coral group has never been simple and it still has many controversial points and ill-defined issues. The abundance and diversity of Faviidae, as well as the broad variability of the morphology of these scleractinian and their skeletal structures, facilitated the appearance of a great number of nominal species with extremely numerous, not always clear or well explained synonymies.
The subdivision of the Scleractinian genus Madrepora, as well as many others, was initiated by Lamarck (1801, 1816), who introduced the following genera: Echinopores, Explanaires, Monticularies, Astrees, Meandrines, belonging at present to the family Faviidae. The first subdivisions of Faviidae, as it was not difficult to understand from their names, were based mainly on growth form of colonies and corallites. Ehinopores and Explanaires had mainly lamellar or folio forms of colonies with well-delineated corallites. Massive and ramose colonies with stellate corallite cones are typical for Monticulaires. Representatives of Astrees have massive colonies with distinct polygonal or round corallites, whereas Meandrines included all corals with corallites, arranged in meandering grooves similar to convolutions of the brain.
Based on the interpretation of Lamarck, two well-defined genera were distinguished later, which were not almost changed afterwards. Echinopora (Dana, 1846) and Monticulaires, later named Hydnophora by Waldheim (1807) were distinguished on the basis of Echinopores and Explanaires. Later on Blainville (1830, 1834) and Dana (1846) followed the same line, and the latter added two genera to Faviidae: Caulastrea and Manicina. The next important step in the development of Faviidae classification was made by Edwards and Haime (1857). Their subdivision of Astrees and Meandrines genera (distinguished by Lamarck) into 8-10 independent genera in many respects anticipated and determined modern views on taxonomy and the number of genera in the Faviidae family. At the same time, they distinguished several groups of related species, attributed to the genus Favites by modern taxonomists, into the independent genera Aphrastraea, Prinastraea and Metastraea. In the opinion of Veron and his co-authors (1977), such intraspecific subdivisions became possible only due to the fact that they were based on a small number of skeletal characteristics of corals, which have been found to be highly variable only recently.
Most of the following investigators took into account the peculiarities of extratentacular division, but they attributed a variety of meanings to this feature. Matthai (1914, 1918) and Crossland (1952) did not consider the nomenclatural status of species groups described by them, but at the same time, they supported differences between species with meandroid and non-meandroidal corallites. Vaughan (1918) held a different view: he raised every of these coral groups up to the family level – Orbicelliidae and Faviidae. Later on Yabe and his co-authors followed Vaughan (Yabe et al. 1936). In the 1940’s Vaughan together with Wells proposed an intermediate point of view. They gave the status of Montastreinae (integrating Ehinopora, Leptastrea, Cyphastrea, Montastrea genera) and Faviidae (Plesiastrea, Favia, Favites, Goniastrea, Hydnophora, Platygyra, Leptoria, Oulophyllia, Caulastrea, Moseleya, Trachyphyllia genera) subfamilies to two groups of Faviidae (genera with extratentacular and genera with intratentacular division in the sense of Edwards and Haime (1857). This statement was widely supported by many researchers: Wells (1956), Nemenzo (1959), Eguchi (1968), Pillai (1972), Wijsman-Best (1972), Pillai and Scheer (1974), and Scheer and Pillai (1974). But Chevalier (1971) came out with an opposite point of view, considering them one group. Vaughan did not distinguish subfamilies within the Faviidae, based on the fact that corals of these genera, reproducing by polyp fission (especially species of Favia genus), exhibit both intra- and extra-tentacular reproduction. Veron and his co-authors (1977) supported this point of view, pointing out that many genera and species have both reproduction modes, one of which usually prevails in one or the other species. Supporting Chevalier’s opinion that the division of the family into of two separate subfamilies was groundless, they list certain species as an example (Favia taxa, Favites pentagona, and Montastrea valenciennesi). They all formally belong to the Faviinae subfamily (i.e. reproduce by intra-corallite division), whereas extratentacular division is also typical for them.
Considering interrelations of taxa within Faviidae, Veron with co-authors noted that a number of genera did not have well-defined limits because of their variation, leaving little or no gap between genera. Generic distinctions are especially difficult in the group of species Favia-Favites-Goniastrea. Discrimination between Favia and Favites is made on the basis of colony morphology, which is placoid for Favia and cerioid for Favites. However, some Favites species (they cite F. rotundata and F. complanata as examples) often display placoid-cerioid colony forms. They write that this fact served as grounds for Matthai (1914, 1918) to unite Favia and Favites under the common name of Favia. stressed Matthai.
The other groups of Faviidae species, in the opinion of Veron and his co-authors, can be subdivided comparatively easily. These are Leptoria, Diploastrea, Cyphastrea, Leptastrea, and Echinopora. On the other hand, they consider that Montastrea and Plesiastrea genera are in the center of this taxonomic debate. One of the reasons for this is the fact that Montastrea has a complicated taxonomic history, including generic names of Orbicella Dana (1846), Heliastrea Edwards and Haime (1857), and Favia Oken (1815) together with some species belonging to them. The other reason was the denial by Vaughan and Wells (1943, Wells, 1956) of the existence of Montastrea species in Indo-Pacific. Because of this, there was confusion not only between these genera, but also between their species, for a long time. It continued until Chevalier (1971) elucidated this problem, explaining that Plesiastrea have real pali, consisting of multitrabecules of an independent diverging structure, and that they together with the obvious absence of direct mesenteries with Plesiastrea polyps serve as important evidence of the difference between the genera Montastrea and Plesiastrea.
When describing Australian Faviidae, Veron and his co-authors (Veron et al. 1977) actually revised all known Faviidae. They re-studied all type material of the valid species, including synonyms, published in taxonomically important works of the 20th century. Following Chevalier (1971), they showed that Faviidae should be united in one Faviidae family without subdivision into taxa of a lower rank. These authors’ point of view on the taxonomy of all Faviidae was partially presented above, but in addition, when describing corals, they expressed their opinion about classification of the most disputed and difficult genera, Favia, Favites, Platygyra, Echinopora, which will be analyzed in detail below. Admitting that Favites are similar to Favia in many respects, but have cerioid colonies usually with septo-thecal septa, they underlined that these distinctions were recognized and taken into account by many researchers, but they were not sufficiently discussed.
The same can be true for Goniastrea, which differ from Favites and Favia, first, by a tendency for corallites to form meandering series, and secondly, purely cerioid corallites have the same calicular structures as meandering ones. There are the primary septa with fine regular denticulation and paliform lobes, forming a prominent crown. At the same time, Favites are characterized by the presence of paliform lobes, whereas septa are presented by a uniform fan-shaped system (Chevalier, 1971). Taxonomic instability within Favites can be explained, first, by arbitrariness of distinctive features of the genus itself in relation to closely-related genera, Goniastrea and Favia in particular, and, secondly, by division of species, in intricate synonymy, especially the series of abdita-halicora-virens-vasta-flexuosa.
Over-devotion to the obsolete nomenclature hierarchy, on the one hand, and insufficient knowledge of variability, on the other hand, existed and still exists to some extent in the taxonomy of the genus Platygyra. Chevalier (1975) described 10 varieties of P. daedalia, 4 varieties of P. sinensis, and divided P. pini, described by him, into two varieties. Logical extension of this process in the presence of a large collection, in the opinion of Veron and his co-authors, will lead to creation of more and more variations, which will result in a situation, when an unlimited nomenclature complex will be created artificially with incomprehensible biological meaning. These authors emphasize in their work that Platygyra consists of relatively few species. Thirty-four nominal specific names were reduced to just four. It was emphasized that they have a broad genotypic and phenotypic variation, and visible variations of skeletal characteristics of corals are fixed within a given species, but their correlation with every other species is too general.
Yuri Latypov, describing the latter Vietnam corals consisting of 17 genera Faviidae, to each of which cast diagnosis and synonymy (Latypov, 1995; Latypov, 2014). He described the 71 species with the synonymy, 40 of them with views of the colonies in situ, and three new to science. Latypov stressed the fundamental differences between Platygyra and Oulophyllia. The second, in contrast to the first deep trough-shaped cups and high walls, developed three cycles of septum, the walls are formed by large exothecal dissepiments. None of these signs is not at Platygyra.
In the last decade, considerable research focuses on the molecular genetic studies and systematics Faviidae (Huang et al., 2009; Budd and Stolarski, 2011; Arrigoni et al., 2012). The purpose of this was not holding pubikatsii taxonomic revision faviid. I tried to analyze the discrepancies and errors in the taxonomic history of these corals and pay attention to the peculiarities of their taxonomic features to facilitate the work on the identification and description of these scleractinian.
2 Morphology and Terminology
2.1 Colonies
Most Faviidae have massive or dome-shaped colonies. Ramose or dendroid projections can be formed in some massive or encrusting-massive colonies. Not many Faviidae can form branched, lamellar or foliose colony shapes. Massive colonies may have polygonal (or angular) corallites, and are called cerioid. Whereas colonies with rounded-cylindrical, disconnected or adjoining corallites are named plocoid. Meandroid colonies, in which corallites are grouped in long winding series similar to the convolutions of the brain, can often be found among the Faviidae (Figure 1, 2).
Presence of an intercorallite structure, formed in the zone of coenosarc-coenosteum, is typical for Faviidae. Coenosteum can be smooth or can consist of exothecal dissepiments, which is referred to as being vesicular. In most Faviidae coenosteum is covered by ribs (costae or coastal ribs) and needle-like spines (spinules), which in turn can be divided or ornamented by many tiny spines and denticles. Distinct development of costae on the exotheca of corallites is one of the most typical features of Faviidae. Costae may be in one and two cycles, equal or unequal, differentiated by size. Costae of neighboring corallites can merge (in such case they are called adjoining) or terminate near the base of the exotheca and be separated by an intercorallite boundary – a narrow crack or groove.
 Figure 1: 1, 3- Favia speciosa, placoid colony with fragmented corallites with well-developed ecsoteca and poorly expressed columella; 2, 4- Favites flexuosa, cerioid colony with polygonal angulate corallites. Total septotheka is clearly visible and adjacent septa, reaching thick columella, different variants of meandroid colonies, 5- Platygyra daedalea, and 6-Oulophyllia crispa. |
 Figure 2: 1- Favia speciosa, 2- Favites flexuosa, principal differences are clearly visible in the form of colonies and corallites and microstructure of their skeletal elements, differences are visible although coverage of cenosarc. |
2.2 Septa
The septal apparatus of the Faviidae are formed by five cycles of septa, often forming a septotheca and, as a rule, differentiated by length into two sizes (orders) with prominent primary septa. Distal septal margins are always more or less dentate serrate with the formation of paliform lobes and pali. External peripheral septal margins are nearly always fan-like and protruding above the theca. The Faviid columella is trabecular, more rarely agminate. It can be lax, consisting of several simple spinules or pinnacles, or very dense, consisting of numerous interlacing trabeculae, often winding spirally. One or several concentric circles, made of merging granulated septal ends, can be formed in a Faviid calice. These circles are called pseudosynapticulae, and almost all of them have a distinct indentation – a depression above the columella.
2.3 Horizontal skeletal elements
Faviidae have rather well developed horizontal skeletal elements: tabulae or dissepiments. Tabulae may be complete or incomplete, convex or concave, rarely with additional lamellae. Dissepiments formed inside the corallite between septa are called endothecal. They are formed by large (up to 5-7 mm), medium (3-5 mm) or small (less than 3 mm) laminae, bulging in various degrees forming vesicles. Dissepimental laminae can be variously oriented within the corallite cavity: with their convexity upwards towards the calice (horizontal), towards the axis (inclined), or towards the corallite axis (vertical). Dissepiments, separating neighboring corallites, often form a vertical row of horseshoe-shaped lamellae. Exothecal dissepiments are formed among (between) corallites and costae. Dissepimental laminae are laid frequently (15-20 per 1 cm of corallite length), moderately frequently (10-15) or rarely (less than 10). They can be thin (less than 0.2 mm) or thick (more than 0.3 mm). Several dissepimental rows, completely filling a corallite cavity from the wall to columella, are called a dissepimentarium (Figure 3).
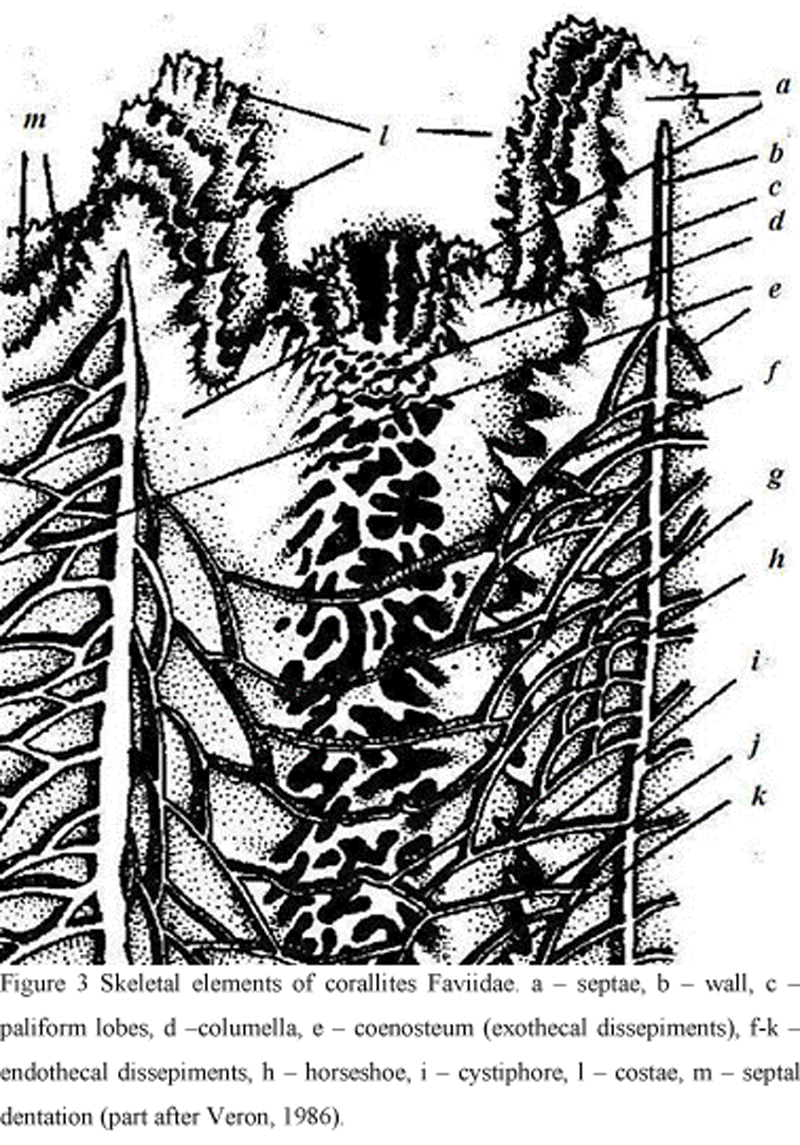 Figure 3 Skeletal elements of corallites Faviidae. a – septae, b – wall, c – paliform lobes, d –columella, e – coenosteum (exothecal dissepiments), f-k – endothecal dissepiments, h – horseshoe, i – cystiphore, l – costae, m – septal dentation (part after Veron, 1986). |
3 Discussion
3.1 Taxonomic peculiarities
As can be seen from the review of Faviidae systematization history, taxonomic problems have existed at higher and lower taxonomic levels. There were different opinions and disputes about the integrity of the Faviidae family and on its value, and on the names of genera included in it. There was no less uncertainty in species identification, as well as at establishing their placement in one or the other genus. I think many researchers can repeat after Chevalier that they are not able to always identify four Echinopora species, and the same is true of other genera. The problem is not much better with the four species of Platygyra. The larger a collection is, the more samples of one species at least from the same area are available, and the more uncertainty and doubts a taxonomist has, since the limits of extreme values of features become more and more extensive, and variability curves become more and more even and continuous.
In principle, this is all right. If these are specimens of the same species, a complex of values of features of all samples must fit in the limits, determined by the given feature, and constitute a complete series of transitive values of the feature. It is all correct, and will work only when the feature is clear and has a single meaning, no matter whether it is qualitative or quantitative. If we limit ourselves to such a terse diagnosis as to say that Favites are similar to Favia but they are cerioid (Wells, 1956), then it will be necessary to follow the diagnosis strictly. Favia have only placoid colonies, and Favites – only cerioid ones, but not subplacoid or subcerioid, as we can often read in diagnoses and descriptions of Faviidae (Figure 1, 2).
3.2 Favia and/or Favites
Strictly speaking, all Favia have placoid colonies, especially if we mean type specimens. In other words, all these corals have massive colonies with corallites closely or distantly located from each other (Figure 1, Figure 2). I stress that these are detached corallites, mainly of a round shape, though this roundness can be close to a trigonal or irregularly polygonal form (F. laxa, F. speciosa). The opinion about purely cerioid Favia pallida and F. rotumana (Veron et al. 1977) is not quite correct. Type specimens of these species are typically placoid. They have round corallites, slightly detached or adjoining. Every corallite has clearly developed endo- and exotheca. Adjoining corallites are necessarily separated by a distinct groove. The same can be said of the majority of cited pictures of these species (Hoffmeister, 1925; Wells, 1954; Chevalier, 1071; Scheerr and Pillai, 1974, 1983; Veron, 1986; Nakamori, 1986; etc.). Even in supposedly subcerioid specimens (Veron et al. 1977, p. 34, Figure 49-52; Figure 62-63) endo- and exotheca (despite the irregular, almost polygonal form of corallites) and clearly detached corals with discrete costal ridges are clearly seen. As to the truly cerioid specimen of F. pallida (p. 34, Figure 46, in the same work), its attribution to the genus Favia seems to me doubtful, as it has typically cerioid corallites with a common endotheca, here and there turning into septotheca, septa of the neighboring corallites are adjoining, and these are features typical for Favites.
Regarding cerioid-placoid forms of Favites rotundata and Favites complanata (Veron et al. 1977), it is necessary to note approximately the same circumstances as discussed above. First, Favites rotundata has now been placed in the genus Favia, secondly, the type specimen and cited pictures of Favites rotundata specimens have clearly polygonal (cerioid) corallites with a common septothecal wall and adjoining septa. A false boundary between corals can be formed at the expense of light removal towards adjoining septa at their transition to a neighboring corallite and replacement of an earlier cycle by a later one. And finally, the columella is formed by complexly divided anastomosing trabeculae, which cannot be in Favia.
In conclusion, we shall consider one more example, testifying to the necessity of a strict approach to the character of a “placoid coral”, which is attributed to the genus Favites by Veron and his co-authors (1977). In this case, first of all, not only the purely placoid form of a colony and corallites alerts one to the problem, but also two cycles of highly projecting septa with a very well developed crown of pali. These features are more typical for the genus Plesiastrea. In addition, the total number of septa (the third cycle is always rudimentary) rarely exceeds 30. In the smallest representative of Favia the number of septa is 30-40 (the third and fourth cycles are always developed), and on the average they have 40-60 septa.
Thus, in many cases taxonomic problems in identification and systematization of Faviidae arise in the result of vagueness of definition of diagnostic classification characters or introduction of alien characters, not proper for the given genus. Because of this process, an artificial extension of natural limits of species variability takes place, with species falling out through the boundaries fixed for each genus.
3.3 Transition some features
In support of the above-said, it is necessary to consider the similarity of growth forms of some species groups (Caulastrea-Favia-Favites- Goniastrea-Platygyra Goniastrea-Platygyra), which was spoken about in the previous chapter when discussing variability and the transition of many features from one genus to the other one. Now we shall see to what extent a two-way transition of such features as “placoid”, “cerioid” and meandroid” growth forms in the series Caulastrea-Favia-Favites-Goniastrea-Platygyra is real. All Caulastrea have only phaceloid colonies, and however close to each other corallites may be located, each of them has epitheca and its colony has no coenosteum. And it must be so, proceeding from the definition of a phaceloid colony (Wells, 1956). In a placoid colony all corallites have a common coenosteum, and phaceloid colonies are not known among Favia. This means that in this case an indicator between Caulastrea and Favia, showing transition of features of colony forms, does not work in both directions. As was shown above, placoid colonies fundamentally differ from cerioid ones. The former have round discrete corallites with endo- and exotheca developed on a smooth or vesicular coenosteum. The latter are characterized by polygonal corallites with a common exotheca, adjoining septa and the absence of coenosteum. This is why a strict approach to the definitions of “placoid” and “cerioid,” taking into account that all Favia are placoid, and all Favites are cerioid, goes a long way toward removing the problem of transition of these features from one genus to the other one. Among Favites the species F. bennettae, having a tendency for corallites to form meanders, is now attributed to the genus Oulophyllia (Veron, 1986), thus, the transition of the feature “meandroid colonies” to the genus Goniastrea and back to Favites is removed.
For the remaining three genera the feature “meandroid colonies” is not really the most reliable character. They all have cerioid and meandroid corallites in some of their species. Goniastrea in most cases are cerioid, Platygyra are both, and Oulophyllia are meandroid with individual polygonal corallites. It means that this feature cannot serve as a diagnostic feature. But, if a characteristic of cerioid-meandroid colonies were added that “well formed large pali, forming a crown, always projecting over the corallite indentation,” then these features would be typical only for the genus Goniastrea. Neither Platygyra nor Oulophyllia have a distinctly developed crown. The spongy columella of Platygyra merges into one line, passing through all corallites within a valley. In Oulophyllia the columella always designates the centers of corallites. In addition, broad trough-shaped valleys or corallites of the Oulophyllia are 1.5 times wider than the funnel-shaped valleys of Platygyra.
3.4 Placoid or cerioid
The characteristics “massive placoid” and “massive-cerioid” are as insufficient for the identification of Favia and Favites without mentioning that they are Faviidae as the characteristic “massive cerioidmeandroid” is insufficient for identification of Goniastrea, Platygyra and Oulophyllia. As it is known from the principles of zoological systematization, a character or a sum of characters become taxonomic characters only in relation between two taxa. And when they are unambiguously limited qualitatively or quantitatively, then specimens of a species will be identified easier, and there will always have to be a gap in the values of one or more characters between different species, no matter whether the characters are morphological, ecological, biochemical or genetic.
Difficulties in identification, and especially in the separation of Favia speciosa from F. pallida, have been discussed repeatedly (Chevalier, 1971, 1975; Veron et al. 1977; Scheer and Pillai, 1983). Now we shall analyze a complex of characters of both species (Table 1). As can be seen, there are distinctions for every character. At the same time, many characters will overlap even though a sample of colonies of the two species is not large. But F. pallida has no highly prominent corallites, and F. speciosa has no corallites joined together. There are also characters with a larger gap. Horizontal skeletal elements of these species corals differ greatly. The difference in lamina size is not very great, but the rates that laminae are laid down in the process of corallite growth differ substantially. In F. speciosa dissepiments are laid 2-3 times more often than in F. pallida. It seems that this genetically determined character is functionally connected and morphologically developed in highly prominent corallites, which are absent in F. pallida corals, having different rates of vertical growth.
 Table 1 Comparison of attributes of two species of Faviidae. |
Good examples of overlapping of values of skeletal characters of various species, and at the same time of their recognition, were cited for the genus Platygyra (Veron et al. 1977). Partially overlapping of characters values can be clearly seen, but on the other hand, in every species its inherent characters of wall width, corallite length and width dominate (Figure 4). Such comparisons and construction of diagrams and graphics based on the data give good results for any characters of all coral groups, both recent and fossil (Veron and Pichon, 1980; Tesakov, 1978; Latypov, 1984, 2014).
 Figure 4 Comparison of the mean valley length (top), mean valley width, and mean wall width per corallum of the Platygyra species indicated. All sizes in mm, on an axis of ordinates number of corolla (part after Veron et al., 1977). |
4 Conclusion
All of the above can also be attributed to classification of the Faviidae. It is necessary to add, however, that the shape of the Fungiidae can be a good generic taxonomic character, as well as circumoral or peripheral budding. The length of fossa, especially its ratio to the coral diameter, can be used as an identification (recognition) character for species. Perforation of septa in various degrees or its absence can be used both when distinguishing taxa at the generic level, and when distinguishing different species within the same genus. The arrangement of secondary stomata in one line or in concentric circles, or its absence is a generic level character. Shape and sizes of septal denticles in accordance with sizes and shape of costal spines serve as good interspecies distinction characters.
In conclusion it is necessary to say that both (ambiguity) of characters with many values leads to an unnatural increase in their variability and the boundaries of taxa, and a clearly determined character very definitely limits its variability within a taxon and allows us to find gaps between taxa.
Arrigoni R, Stefani F, Pichon M, Galli P, Benzoni F., 2012 Molecular phylogeny of the robust clade (Faviidae, Mussidae, Merulinidae, and Pectiniidae): an Indian Ocean perspective. Molecular Phylogenetics and Evolution. Oct 31; 65 (1):183-93.
Budd AF, Stolarski J. Corallite wall and septal microstructure in scleractinian reef corals: comparison of molecular clades within the family Faviidae. Journal of Morphology. 2011 Jan 1; 272 (1):66-88.
http://dx.doi.org/10.1002/jmor.10899
Blainville H.M., 1830, Zoophytes. Ibid, 60: 310-358
Blainville H.M., 1834, Manuel d'Actinologie ou de Zoophytologie. 2(8): 694 p
Chevalier J.P., 1971, Les Scléractiniaires de la Mélanésie franç aise (Nouvelle Calédonie, îles Chesterfild, îles Loyauté, Nouvelles Hebrides). Pt 1, Ibid, 5: 5-307
Chevalier J.P., 1975, Les Scléractiniaires de la Mélanésie française (Nouvelle Calédonie, îles Chesterfild, îles Loyauté, Nouvelles Hebrides). Pt 2, Ibid, 7: 5-7
Crossland C., 1952, Madreporaria, Hydrocorallinae, Heliopora and Tubipora // Sei. Rep. Great Barrier Reef Exped. 1928-1929. L.: British Museum (Natural History). 6: 85-257
Dana J.D., 1846-1849, Zoophytes, US Exploring Exped., 1838-1842. New York. 7: 1-740
Edwards H.M., Haime J., 1857.Histoire naturelle des Coralliaires. P., 1: 326 p
Eguchi M., 1968, The hydrocorals and scleractinian corals of Sagamy Bay. Tokyo: Aruzen, C1-C74
Forsckål P., 1775, Descriptions Animalium, Avium, Amphibiorum, Piscium, Insectorum, Vermium que in itinere orientali observavit Petrus Forsckål Mölleri, Haunia, 164 p.
Hoffmeister J.E., 1925, Some corals from American Samoa and the Fiji Islands // Pap. Dep. Mar. Biology Carnegie Institution Washington. 22: 1-90
Huang D, Meier R, Todd PA, Chou LM. More evidence for pervasive paraphyly in scleractinian corals: Systematic study of Southeast Asian Faviidae (Cnidaria; Scleractinia) based on molecular and morphological data. Molecular Phylogenetics and Evolution. 2009 Jan 31; 50 (1):102-16.
http://dx.doi.org/10.1016/j.ympev.2008.10.012
Lamarck J.B. P.A. de M., 1801, Système des animaux sans vertebres // P.: Deterville, 432 p
Lamarck J.B. P.A. de M. 1815-1816, Histoire naturelle des animaux sans vertèbres, P.: Verdière. 2:. 568 p
Latypov Yu.Ya., 1995, Scleractinian corals of Vietnam. Part III. Faviidae. Fungiidae. Moscow: Nauka, 144 p.
Latypov Yu.Ya., 1984, Drevneishie odinochnye korally i princypy ih systematiki. M.: Nauka. 92 p, (In Russian)
Latypov Yu.Ya., 2012, Encrusting protected reef Hon Nai in Cam Ranh Bay in the South China Sea, Natural Science. 4: 14–21
http://dx.doi.org/10.4236/ns.2012.41003
Latypov Yu.Ya., 2013, Favia camranensis sp. n. (Scleractinia: Faviidae), a New Coral Species from Southern Vietnam, Russian Journal of Marine Biology, 39(3): 223–224 Latypov Yuri Ya., 2014, Scleractinian corals of Vietnam Science Pyblishing group.. pp. I-XXXIV, 1-481
Matthai G., 1914, A revision of the recent colonial Astraeidae possessing distinct corallites. Transaction of the Society of London. Ser. 2, Zool. 17: 1-140
Matthai G., 1948, Madréporaires de Nouvelle-Calédonie Bulletin biologique de la France et de la Belgique. 597: 70-88
Nakamori T., 1986, Community structures of Recent and Pleistocene hermatypic corals in the Ryukyu Islands, Japan Sei. Rep. Tohoku Univ. Sendai. Ser. 2, Geol. 56: 71-133
Nemenzo F., 1959, Systematic studies on Philippine shallow water Scleractinians. 2. Suborder Faviidae Nat. Appl. Sei. Bull. Philippines. 16(1/4): 73-135
Oken, L. 1815. Lehrbuch der Naturgeschichte. HI. Zoologie. Leipzig; Jena 1: 59-74.
Pillai C.S.G., 1972, Stony corals of the sea around India, Proc. Symp. Corals and Coral Reefs, 1969 Marine Biologycal Association India. Delhi, 191-216
Pillai C.S.G., Scheer G., 1974, On a collection of Scleractinia from the Strait of Malacca, Proc. II Intern. Coral Reef Symp. Brisbane, 1: 445-464
Scheer G.. Pillai C.S.G., 1974, Report on Scleractinia from the Nicobar Islands II, Zoologica. 42(122): 1-75
Scheer G., Pillai C.S.G., 1983, Report on the stony corals from the Red Sea, Ibid. 45(133): 1-98
Tesakov Yu.I., 1978, Intraspecific divisions of Tabulata to their variability from positions of the biological concept of a species. In Drevnie CNIDARIA, Novosibirsk: Nauka, 128-131. (In Russian)
Vaughan T.W., 1918, Some shoal-water corals from Murray Islands, Cocos-Keeling Islands and Fanning Islands Pap. Dep. Mar. Biol. Carnegie Inst. Wash. 9(213): 51-234
Vaughan T.W., Wells J.W., 1943, Revision of the sub-order, families and genera of the Scleractinia Geol. Soc. Amer. Spec. Pap. 44: 506-510
Veron J.E.N., 1986, Corals of Australia and the Indo-Pacific. North Ryde (N.S.W.): Angus and Robertson. 644 p
Veron, J.E.N., Pichon M., 1980, Scleractinia of Eastern Australia. Part 3. Families Agaraciidae, Siderastreidae, Fungiidae, Oculinidae, Merulinidae, Mussidae, Pectiniidae, Caryophylliidae, Dendrophylliidae. Austral. Inst. Mar. Sci. Monogr., IV(1): 471 p
Veron J.E.N., Pichon M., Best M.W., 1977, Scleractinia of Eastern Australia Austral. Inst. Mar. Sci. Monogr. Ser. 3: 1-233
Waldheim von F.G., 1807, Museum Demidoff, ou,Catalogue systématique et raisonné des curiosités de la nature et de l’art: données à l’Université Impériale de Moscou par son excellence Monsieur Paul de Demidoff, Moscow: Imprimerie de Université Impériale de Moscou. 3: 300 pp., 6 pls
Wells J.W., 1954.Recent corals of the Marshall Islands US Geol. Surv. Prof. Pap. 260-1: 385-486
Wells J.W., 1956.Scleractinia. Treatise on invertebrate paleontology, Lawrense, Pt F: Coelenterata. 328-440
Wijsman-Best M., 1972, Systematics and ecology of New Caledonia Faviinae (Colenterata, Scleractinia), Bijdr. Dierkd. 42(1): 1-76
Yabe H., Sugiyama T., Eguchi M., 1936. Recent reef-building corals from Japan and the South Sea Islands under the Japanese mandate. Ibid. Spec. 1: 1-66



. PDF(563KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Yuri Latypov
Related articles
. Faviidae
. History
. Taxonomy
. Signs
Tools
. Email to a friend
. Post a comment