Salt-induced Modifications in the Vegetative Anatomy of Bottonweed and Peruvian Spikesedge 
 Author
Author  Correspondence author
Correspondence author
International Journal of Marine Science, 2016, Vol. 6, No. 47 doi: 10.5376/ijms.2016.06.0047
Received: 28 May, 2016 Accepted: 08 Aug., 2016 Published: 08 Aug., 2016
Kekere O., 2016, Salt-induced Modifications in the Vegetative Anatomy of Bottonweed and Peruvian Spikesedge, International Journal of Marine Science, 6(47): 1-12 (doi: 10.5376/ijms.2016.06.0047)
Leaf, stem and root anatomy of bottonweed (Diodia maritima Thonn.) and Peruvian spikesedge (Kyllinga peruviana Lam.) irrigated with 0 (control), 70, 140, 280 and 560 mM NaCl solutions were examined under the microscope in a screen house study at Adekunle Ajasin University, Akungba Akoko, Ondo State, Nigeria. D. maritima did not survive beyond 280 mM and stomata were present only on the abaxial leaf surface in both plants. Salinity decreased leaf stomatal density significantly in D. maritima but increased it at 70-140 mM and sharply decreased it at 280-560 mM in K. peruviana. The number of stomata per leaf declined significantly at 140-280 mM in D. maritima but increased significantly at all salt levels in K. peruviana. Spongy cell proportion increased under salt in the plants but significant only in K. peruviana. Palisade and spongy cell diameters were improved by salt treatment with a significant difference at 140-280 mM in D. maritima and at all salinity levels in K. peruviana. Salinity increased stem diameter, percentage stele, pith and cross sectional area significantly at 70-140 mM in D. maritima but decreased the parameters in K. peruviana except at 280 mM. In the root, the diameter, cross sectional area and percentage cortex were reduced in D. maritima but increased in K. peruviana under salt treatment except at 560 mM for stem diameter and at 280-560 mM for cross sectional area and percentage cortex. The anatomical modifications varied considerably in strand plants with implications on their adaptation for survival in the strandline.
1 Introduction
Strandlines are created along the high water mark on a range of habitats including coastal vegetated shingle, sand dunes and salt marshes (Lee and Ignaciuk, 1985). They are coastal environments above high water, which is out of reach of the average high tide and it is characterized by the deposition of drift materials, litter, debris and many discarded items left behind by the previous receding tide (Rozema et al., 1982). Plate A shows a view of a strandline environment in Nigeria. They have worldwide distribution with ecological, social and economic values (Williamson, 2005). The vegetation provides essential habitat, spawning sites and food for many diverse species of terrestrial and marine wildlife. They are able to withstand periodic coastal disturbances and they are important on exposed shores, where they act as precursors to sand dunes (Williamson, 2005), as shown in Plate B.
Strandline environment is usually occupied by only few plant species due to the severity of the environment (Rozema et al., 1982) as several environmental factors affect the vegetation, and salinity is possibly the most imperative ecological restriction that determines the species existence (Owen et al., 2001; Peter et al., 2003; Pichler and Oberhuber, 2007), thus plant ability to survive in the habitat is greatly determined by its adaptation to salinity.
Most studies on salinity tolerance in strand plants have focused mainly on morphological and physiological adaptations (Barbour et al., 1985; Flowers and Colmer, 2008) while anatomical adaptations which can also offer some explanations of species abilities to grow on the strandlines, and can be easily recognized and quantified have been largely ignored.
 Plate A A view of the strandline environment along Arogbo Ijaw Seaside Beach, Ondo State, Nigeria |
.png) Plate B A view of some plant species at Arogbo Ijaw Seaside Beach, Ondo State, Nigeria Note: DI= discarded item by tide |
It is evident that there are big changes in anatomy of plants growing in saline soils (Ediga et al., 2013). Depending on the species, anatomical changes in plants may be either adaptations to salinity stress or signs of damage and disturbance of the normal life processes (Yeo and Flowers, 1980). Salinity affects cell division and expansion processes, reduces the size of apical meristems, cortex and vascular cylinder (Zidan et al., 1990). Also, salinity causes changes in tracheary element density, lumen volume and vessels diameter (Junghans et al., 2006; Escalante-Perez et al., 2009; Chen and Polle, 2010) as well as stomatal density, shape, and size (Abbruzzese et al., 2009). Anatomical changes can affect plant ability to conduct water and nutrients, thus anatomical adaptations are as important as morphological and physiological processes (Younis et al., 2014). Some specific anatomical adaptations to salinity stress include increased succulence by cell expansion, changed number and size of stomata, thickened cuticle, inhibited differentiation and changed diameter and number of xylem vessels, development of tyloses and earlier occurrence of casparian strips and lignification (Flowers et al., 1986).
 Plate C Photograph of Diodia maritime |
 Plate D Kyllinga peruviana growing on the strandline along Nigerian coastline |
Buttonweed (Diodia maritima Thonn.) belongs to the family Rubiaceae. It is a perennial prostrate herb usually rooting at nodes, rarely scandent or somewhat tufted. The leaves are ovate and subacute at apex, cuneate into a short petiole at base and not fleshy and glabrous (Hutchinson et al., 1968) as shown in Plate C. Peruvian spikesedge (Kyllinga peruviana Lam.) belonging to the Cyperaceae family. It is rhizomatous sedge with a distinctive spherical, white inflorescence that is densely crowed with very small greenish yellow flowers. The flowers are in spikes or spikelets and the perianth is either reduced or represents only a set of bristles or scales. It often has leafless stems and the leaves are sword shaped, narrow and flat. It propagates itself mainly by the rapid growth of its spreading rhizome as shown in Plate D. Both plants are widely distributed and restricted to strandline habitat in Nigeria where they grow naturally in a short distance from sea on the sandy beaches (Hutchinson et al., 1968).
Both plants were chosen for this study because they are among the few species found growing naturally, abundant, wide spread and restricted to the habitat. I hypothesized that they have some structural anatomical changes for survival under salt stress, which are largely unavailable in scientific literatures. This research is therefore aimed at investigating leaf, stem and root anatomical changes in the coastal halophytes, Diodia maritima and Kyllinga peruviana under salinity stress in order to boost scientific knowledge on some of the adaptation mechanisms of salinity tolerance in strand plants, which will be useful in breeding program to evolve salt-tolerant crops.
2 Materials and Method
2.1 Plant materials
Young rhizomes of Kyllinga peruviana and seedlings of Diodia maritima collected from the coastal sand dune at Arogbo Ijaw Seaside Beach, Ondo State, Nigeria (60 111N latitude, 40 291E Longitude) were used as planting materials. Uniform plants raised from the young rhizomes of Kyllinga peruviana and seedlings of Diodia maritima were grown in individual perforated plastic pots (20 x 26 cm) filled with 2:1 mixture (v/v) of river sand to topsoil.
2.2 Planting soil
The soil used for planting was a 2:1 mixture (v/v) of river sand to topsoil. The sand was collected from river sand deposited at a construction site while top soil was collected from the experimental farm of Plant Science and Biotechnology Department, both at Adekunle Ajasin University, Ondo State, Nigeria (Latitude 70 28’N, Longitude 50 44’ E). Both soils were air-dried, thoroughly mixed at 2:1 mixture (v/v) and filled into the perforated plastic pots. The soil was passed through a 2-mmol/L sieve, and analyzed for the physico-chemical parameters. Particle distribution of the soil was obtained using the rapid method, while pH was measured in 1:1 soil: water suspension. Nitrogen was determined by the modified Kjeldahl method while phosphorus was assayed by Bray’s P1 solution and read on a spectrophotometer. Cations were extracted with 1.0 M acetate solution at pH 7.0; sodium and potassium contents in the extract were determined by flame photometry while calcium and magnesium were obtained by atomic absorption spectrophotometry. Organic carbon was determined by the wet oxidation method while cation exchange capacity (CEC) was by distillation method particle distribution using the rapid method Soil pH was measured in 1:1 soil: water suspension. Nitrogen was determined by the modified Kjeldahl method while phosphorus was assayed by Bray’s P1 solution and read on a spectrophotometer. Cations were extracted with 1.0 M acetate solution at pH 7.0; sodium and potassium contents in the extract were determined by flame photometry while calcium and magnesium were obtained by atomic absorption spectrophotometry. Organic carbon was determined by the wet oxidation method while cation exchange capacity (CEC) was by distillation method, following the standard method of the Association of Official Analytical Chemists (AOAC, 1985) at the Central Laboratory of National Institute for Oil Palm Research (NIFOR), Nigeria. The soil was a sandy soil with physico-chemical parameters of 5.48 pH, 20.42 ppm N, 3.56 ppm P, 3.56 (meg/100g) K, 2.32 (meg/100g) Ca, 2.60 (meg/100g) Mg, 8.2 (meg/100g) CEC, 3.67% C, 80.68% sand, 12.06% silt and 8.36% clay (Kekere et al., 2016).
2.3 Preparation of saline solution
Concentrations of 70 mmol/L, 140 mmol/L, 280 mmol/L and 560 mmol/L NaCl saline solutions were prepared in plastic kegs just before each treatment by dissolving weighed amount of available salt in tap water to make the desired concentrations. One molar solution was prepared by dissolving 58.5 g (molar mass of NaCl) in water to make 1 litre. This quantity (58.5 g) was divided by 1000 to make 1 mmol/L solution, and was subsequently multiplied by the desired value of mmol/L and number of litres to be prepared, to account for the amount of salt to be dissolved in water.
Plants were irrigated 3 times/week with saline water at concentrations of 0 (control), 70 mmol/L, 140 mmol/L, 280 mmol/L and 560 mmol/L mmol/L NaCl. Saline water treatment was initiated by gradual 70 mmol/L increments at 2-day intervals to reach the maximum salinity level of 560 mmol/L NaCl at 14 days, to prevent osmotic chock. Irrigation was done at the root zone of the plant to ensure that the relative level of soil salinity would be the primary cause of any observed effect rather than combined effect of soil and air-borne salinity. Each pot was treated with 150 ml (volume enough to keep the soil moist) per treatment; thus each pot received 450 ml of NaCl solution per week. The treatments lasted for 12 weeks; thus, each pot received a total of 4.5 litres of saline solution before the experiment was terminated. Meanwhile, pots were saturated with water and allowed to drain once per week to flush out excess salt and prevent salt build-up in the soil above the treatment levels. Besides, salt concentration in the pots was monitored by conductivity measurement in the leaked excess solution after application. The experiment was carried out from August to November, 2015. It was completely randomized with 5 single-plant replicates per treatment.
2.4 Anatomical studies
After 12 weeks of saline water irrigation, stomata number was estimated on the leaves sectioned for anatomical studies following the method earlier described by Hwang and Chen (1995). The stomata number per leaf was estimated as the product of stomata density and leaf area, measured with leaf area meter (LI-COR 300 model). Following the conventional method of free hand sections (Longstreth and Nobel, 1979), samples were taken from the centrally located region at each side of the mid-vein of the youngest fully expanded leaf. Stem sections were made at 5 cm from the tip of the apical bud based on the standard freehand section method (Ruzin, 1989). Segments from the root absorption zone (1.5 cm from the tip) were sampled. Unstained fresh free hand sections were cut in series, infiltrated with distilled water and examined. Measurements and micrographs were made using a digital camera (Nikon FDX-35) equipped with a Nikon stereo-microscope (Nikon 104, Japan). In leaf, the palisade cell length and diameter, spongy cell diameter, as well as the proportions of palisade and spongy cells expressed as a percentage of leaf thickness were measured. In stem and root, their diameter, cortex, pith and stele zone thickness expressed as a percentage of the cross section were determined.
2.5 Statistical analysis
Data were subjected to single factor analysis of variance and means were separated with Turkey Honest Significant Difference (HSD) test using SPSS version 17.0 (SPSS Inc., Chicago, IL, USA) at 95% level of significance.
3 Results
Diodia maritima leaf stomatal density decreased significantly with increasing salt concentration (Table 1). It reduced by 33.01%, 58.62% and 81.90% at 70 mmol/L 140 mmol/L and 280 mmol/L NaCl respectively relative to the control. In Kyllinga peruviana however, leaf stomatal density increased over the control by 24.35% and 21.61% at 70 mmol/L and 140 mmol/L respectively, but sharply decreased by 15.96% and 39.17% at 280 mmol/L and 560 mmol/L concentrations respectively (Table 1). The values of the number of stomata per leaf declined significantly in both plant species with increasing concentration of salt (Table 1, Plates E and F).
In Diodia maritima, the decline became significant at concentrations of 140 mmol/L and 280 mmol/L with 73.88% and 92.98 % reduction respectively. On the other hand, K. peruviana had significantly reduced the number of stomata per leaf at all levels of salt treatment relative to the control, with percentages of 25.81, 31.92, 57.33 and 76.42 at 70 mmol/L, 140 mmol/L 280 mmol/L and 560 mmol/L concentrations of NaCl respectively. Plants grown under salt application had thicker leaves than those irrigated with salt-free water (Table 2).
However, increase in leaf thickness became significant at 140 mmol/L and 280 mmol/L in Diodia maritima whereas it was significantly higher at all levels of salinity over the control in Kyllinga peruviana. The proportion of spongy cell per leaf also increased in both plants but there was no significant difference between the control and those exposed to salt stress in Diodia maritima while significantly higher values were obtained in Kyllinga peruviana subjected to saline water irrigation at all levels than in the control. In addition, one-way ANOVA showed that the palisade cell length was not affected by salt application. Furthermore, the palisade and spongy cell diameters had similar trends in both plant species. The parameters were improved by salt treatments in both plants. While Diodia maritima showed a significant difference at 140 mmol/L and 280 mmol/L only, Kyllinga peruviana gave significantly higher values at all salinity levels when compared to the control. Salinity increased stem diameter, percentage stele, pith and cross sectional area significantly at concentrations of 70 mmol/L and 140 mmol/L NaCl but without statistical difference at 280 mmol/L in Diodia maritima (Table 3, Plate G).
.png) Table 1 Effect of salinity on leaf stomatal density and number of stomata per leaf on the abaxial leaf surface of Diodia maritime and Kyllinga peruviana |
 Plate E Effect of salinity on the leaf stomata number of Diodia maritim Note: S = stomata, Magnification= 10 x 10 |
The percentage cortex was also not significantly affected at 280 mmol/L by salt treatment, but it was significantly reduced at 70 mmol/L and 140 mmol/L NaCl. In the root, the dimension of diameter, percentage cortex and cross section area declined significantly under salinity stress compared to the control whereas the proportion of stele was not affected significantly. In K. peruviana stem, the diameter, cross sectional area and proportions of stele and pith had higher values in plants subjected to salinity treatment than in those irrigated with salt-free water (Table 4, Plate H).
4 Discussion
Stomatal density and number of stomata reduction under salinity in D. maritima and at 560 mmol/L NaCl in K. peruviana conforms to the earlier findings in Alternanthera maritima and erecta subsp. maritima where salinity reduced stomata density and number of stomata per leaf (Kekere et al., 2016). Salinity has also caused changes in stomatal density, shape, and size in Populus euphratica (Chen et al., 2015) and Kandelia candel (Hwang and Chen, 1995). Salinity resulted in increased stomatal density at 50 mmol/L, 100 mmol/L, and 150 mmol/L NaCl but decreased at 200 mmol/L salt concentration in Populus euphratica (Alam et al., 2014), similar to what was obtained in K. peruviana in this study. Reduction in stomata is an adaptation to survive salt-stress-induced water deficit by minimizing loss of water through transpiration (Flowers et al., 1986). Salinity led to changes in the leaf cellular structure of the plants. The increase in leaf thickness in this study has similarity with the report on Leucaena leucocephala with increased length and diameter of leaf palisade cells and reduction in intercellular spaces (El-Lamey, 2015). The enlargement of palisade and spongy cells in the plants confirms a high ratio of mesophyll area/leaf area, which has been linked to increment in photosynthetic rate in plants (El-Lamey, 2015). In thick leaves, more mesophyll cells increase internal leaf surface area for gaseous exchange, which improves carbondioxide uptake, thus enhancing photosynthetic rates (Hwang and Chen, 1995).
.png) Table 2 Effect of salinity on the leaf thickness, ratio of spongy and palisade parenchyma to leaf thickness, and mesophyll cell dimensions in the leaves of Diodia maritime and Kyllinga peruviana |
.png) Table 3 Diameter, cross sectional area and proportion of cortex, stele and pith to cross section in the stem and root of Diodia maritima grown in soil with different concentrations of NaCl |
 Plate F Stem anatomy of Diodia maritima under different levels of salinity Note: E= Epidermis, C= Corte, S = stele, P = pith. Magnification = 10 x 40 |
 Plate G Stem anatomy of Diodia maritima under different levels of salinity Note: E= Epidermis, C= Cortex, P= pith, S = stele. Magnification = 10 x 40 |
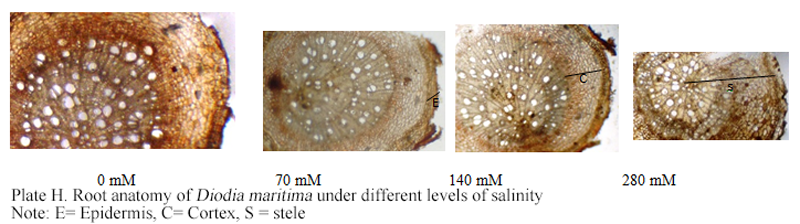 Plate H Root anatomy of Diodia maritima under different levels of salinity Note: E= Epidermis, C= Cortex, S = stele |
|
Table 4 Diameter, cross sectional area and proportion of cortex, stele and pith to cross section in the stem and root of Kyllinga peruviana grown in soil with different concentrations of NaCl |
.png)
The vascular bundle being an integral part of the stele, the changes in the dimension of the stele is an evidence that salinity induces structural changes in xylem and phloem vessels. The improvement in stele proportion due to salinity in this study agrees with the earlier results obtained on jatropha plants (Abou-Leila et al., 2012) and Alternanthera maritima and erecta subsp. maritima (Kekere et al., 2016).
Salinity increased stem cross-sectional area and diameter in D. maritima and K. peruviana by increasing the percentage stele as influenced by the pith. Salinity usually results in a biochemical loosening of the cell wall under turgor pressure, which initiates cell expansion followed by water and solute uptake, and an increased succulence (Chen et al., 2015). Any increase in cortical area under salt stress in the salt tolerant species may be vital under physiological drought for its better water storage, which is an adaptation for ion dilution to minimize the effect of Na+ and Cl- in plant tissues (El-Lamey, 2015). A reduction in cell size under high level of salt has also been observed in Tagetes patula and Ageratum mexicanum (Zapryanova and Atanassova, 2009). Reduction in cell size was also attributed to the plant ability to reduce its size in order to minimize salt uptake (Zapryanova and Atanassova, 2009). The negative effect of salinity on stem and root cells was occasioned by reduction in the vascular area and cortical cells, which was in line with the results earlier obtained in Gazania harlequin (Younis et al., 2014). The reduction in D. maritima root size by salt stress and at high concentration in K. peruviana agrees with the results on Prosopis tamarugo in which root diameter, cortex and vascular system size declined at NaCl concentration up to 400 mmol/L (Serrato-Valenti et al., 1991). Salt stress usually results in water deficit, which causes cell collapse, thus a reduction in cell size (Junghans et al., 2006; Ficho et al., 2009; Escalante-Perez et al., 2009). The root anatomy of the plants were also affected by salinity. It has earlier been reported that plants roots are potentially the first sites of damage or line of defense due to the direct contact with soil solution and the first to encounter excess salinity (Rewald et al., 2013). Increased dimension in the root size at 70~140 mmol/L NaCl in K. peruviana agrees with the report that the root cortex thickness was promoted under 100~200 mmol/L NaCl in the xerohalophyte shrubs Nitraria retusa and Atriplex halimus (Boughalleb et al., 2009). Root cell decrease was obtained in Portulaca oleracea treated with saline water (Alam et al., 2014) similar to the results on D. maritima at all concentrations and at 560 mmol/L NaCl in K. peruviana. Likewise, the cortical parenchyma in Chloris gayana decreased under saline conditions (C´eccoli et al., 2011), and reductions of cortex thickness under high salinity were also reported in shrubs (Boughalleb et al., 2009) and tree roots, which could be as a result of ion toxicity caused by Na+ and Cl- accumulation in the tissues (Reinoso et al., 2004).
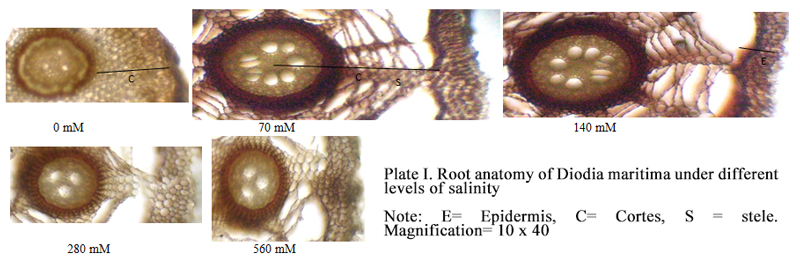 Plate I Root anatomy of Diodia maritima under different levels of salinity Note: E= Epidermis, C= Cortes, S = stele. Magnification= 10 x 40 |
5 Conclusion
Anatomical modifications of the plants to salinity stress depending on the species and salt concentration. However, the results obtained in this study showed an improvement in mesophyll cell size, which usually increases internal leaf surface area for gaseous exchange, hence an improvement in oxygen/carbondioxide uptake, which enhances rates of respiration and photosynthesis in plants. Also, an increase in cell size in stem usually gives better water storage, which is an adaptation for ion dilution to minimize the effect of Na+ and Cl- toxicity. Reduction in the number of stomata per leaf will minimize water loss through transpiration in order to cope with salt-induced water deficit. Reduction in cell size was also attributed to the plant ability to reduce its size in order to minimize salt uptake. Thick root due to inhibition of cell division in the apical meristem of root will minimize salt uptake. Meanwhile, a long-term salinity experiment under field conditions will also better reveal the anatomical modifications of the plants to salt stress.
Abbruzzese G., Beritognolo I., Muleo R., Piazzai M., Sabatti M., Mugnozza G.S. and Kuzminsky E., 2009. Leaf morphological plasticity and stomatal conductance in three Populus alba L. genotypes subjected to salt stress. Environment and Experimental Botany, 66: 381-388
Abou-Leila B., Metwally S.A., Hussen M.M. and Leithy S.Z., 2012. The combined effect of salinity and ascorbic acid on anatomical and physiological aspects of jatropha plants, Australian Journal of Basic and Applied Science, 6(3): 533-541
Alam M.A., Juraimi A.S, Rafii M.Y., and Hamid A.A., 2014, Effect of Salinity on Biomass Yield and Physiological and Stem-Root Anatomical Characteristics of Purslane (Portulaca oleracea L.) Accessions, BioMed Research International, 2015: 1-15
https://doi.org/10.1155/2014/296063
Barbour, M., DeJong, T. M. and Pavlik, B. M., 1985. Marine beach and dune plant societies. In: Physiological ecology of North American plant communities, Chabot, B.F. and Mooney, H. A. (eds.), Chapman and Hall, New York, NY. pp. 296-322
https://doi.org/10.1007/978-94-009-4830-3_14
Boughalleb F., Denden M., and Tiba B.B., 2009, Anatomical changes induced by increasing NaCl salinity in three fodder shrubs, Nitraria retusa, Atriplex halimus and Medicago arborea, Acta Physiologiae Plantarum, 31(5): 947-960
http://dx.doi.org/10.1007/s11738-009-0310-7
Ceccoli G., Ramos J.C., Ortega L.I., Acosta J.M., and Perreta M.G., 2011, Salinity induced anatomical and morphological changes in Chloris gayana Kunth roots, Biocell, 35(1): 9-17
Chen R.Y., and Ayup M., 2015, Effects of high salinity on physiological and anatomical indices during the early stages of Populus euphratica growth, Russian Journal of Plant Physiology, 62 (2): 229-236
http://dx.doi.org/10.1134/S1021443715020168
Chen S.and Polle A., 2010. Salinity tolerance of Populus. Plant Biology, 12:317–333
https://doi.org/10.1111/j.1438-8677.2009.00301.x
Ediga A., Hemalatha S. and Balaji M., 2013. Effect of salinity stress on antioxidant defense system of two finger millet cultivars (Eleusine coracana (L.) Gaertn) differing in their sensitivity. Advanced Biological Research, 7:180-187
El-Lamey T.M., 2015, morphological and anatomical responses of Leucaena leucocephala (Lam.) de Wit. and Prosopis chilensis (Molina) Stuntz to salt conditions, Journal of Applied Biology and Environmental Sciences, 5(7):43-51
Escalante-Perez M., Lautner S., Nehls U., Selle A., Teuber M., Schnitzler J.P., Teichmann T., Fayyaz P., Hartung W., Polle A., Frommol L.J., Hedrich R., and Ache P., 2009, Salt stress affects xylem differentiation of grey poplar (Populus canescens), Plantarium, 229: 299-309
http://dx.doi.org/10.1007/s00425-008-0829-7
Fichot R., Laurans F., Monclus R., Moreau A., Pilate G., and Brignolas F., 2009, Xylem anatomy correlates with gas exchange, water-use efficiency and growth performance under contrasting water regimes: evidence from Populus deltoides x Populus nigra hybrids, Tree Physiology, 29: 1537-1549 PMid:19825869
http://dx.doi.org/10.1093/treephys/tpp087
Flowers T.J. and Colmer T.D., 2008. Salinity tolerance in halophytes. New Phytologist, 179:945–963
https://doi.org/10.1111/j.1469-8137.2008.02531.x
Flowers T.J., Hajibagheri M.A., and Clipson N.J.W., 1986, Halophytes, Quarterly Review of Biology, 61: 313-337
http://dx.doi.org/10.1086/415032
Hutchinson J., Dalziel, J. M. and Hepper, F. N., 1968. Phylogenic sequence of orders and families. In: Flora of West Tropical Africa. Published by Crown Agents for Overseas Governments and Administrations, Volume III, Part 1, Millibank, London, S. W. 1. 574p
Hwang Y., and Chen S., 1995, A natomical responses in Kandelia candel (L) Druce seedlings growing in the presence of different concentrations of NaCl, Bulletin of Botanical Society of Academia Sinica, 36: 181-188
Junghans U., Polle A., Duchting P., Weiler E., Kuhlman B., Gruber F., and Teichmann T., 2006, Adaptation to high salinity in poplar involves changes in xylem anatomy and auxin physiology, Plant Cell Environment, 29: 1519-1531
https://doi.org/10.1111/j.1365-3040.2006.01529.x
Kekere O., Bulu Y.I., and Ologundudu F., 2016, Anatomical adaptations in beach alternanthera and white mouth dayflower under salinity stress, Plant Cell Biotechnology and Molecular Biology, 17(5&6):213-222
Lee J A and Ignaciuk R., 1985. The physiological ecology of strandline plants. Plant Ecology. 1985; 62 (1-3): 15-19
https://doi.org/10.1007/978-94-009-5524-0_36
Longstreth D. J. and Nobel P. S., 1979. Salinity effects on leaf anatomy: Consequences for photosynthesis. Plant Physiology, 63: 700-703
https://doi.org/10.1104/pp.63.4.700
Peter, C. I., Ripley, B. S. and Robertson, M. P., 2003. The distribution of Scaevola plumieri along the South African coast is limited by seasonal water balance and temperature. Journal of Vegetation Science, 14: 89-98
Pichler, P. and Oberhuber, W., 2007. Radial growth response of coniferous forest trees in an inner Alpine environment to heat-wave in 2003. Forest Ecology and Management, 242: 688–699
https://doi.org/10.1016/j.foreco.2007.02.007
Reinoso H., Sosa L., Ram´ırez L., and Luna V., 2004, Salt-induced changes in the vegetative anatomy of Prosopis strombulifera (Leguminosae), Canadian Journal of Botany, 82(5):618-628
http://dx.doi.org/10.1139/b04-040
Rewald B., Shelef O., Ephrath J. E., and Rachmilevitch S., 2013, Adaptive plasticity of salt-stressed root systems, In: Ecophysiology and Responses of Plants under Salt Stress, Ahmad P., Azooz M.M., and Prasad M.N.V., Eds., chapter 6, pp. 169-201, Springer, New York, USA
http://dx.doi.org/10.1007/978-1-4614-4747-4_6
Rozema, J., Bijl, F., Dueck, T. and Wesselman, H., 1982. Salt-spray stimulated growth in strand-line species. Physiologia Plantarum, 56: 204-210
https://doi.org/10.1111/j.1399-3054.1982.tb00326.x
Ruzin, S. E., 1989, Plant micro-technique and microscopy. Oxford University Press, Inc., New York
Serrato Valenti, G., Ferro, M., Ferraro, D. and Riveros, F., 1991, Anatomical changes in Prosopis tamarugo Phil. Seedlings growing in different levels of salinity. Annals of Botany, 68: 47-53
Williamson, K. (2005). Action plan scope. In: Strandlines. Nature Gwynedd Bulletin, version 1, 2005. 6p
Yeo, A. R. and Flowers, T. J., 1980, Salt tolerance in the halophyte Suaeda maritima L. Dum.: Evaluation of the effect of salinity upon growth. Journal of Experimental Botany, 31 (4): 1171-1183
https://doi.org/10.1093/jxb/31.4.1171
Younis A., Riaz A., Ahmed I., Siddique M. I., Tariq U., Hameed M., and Nadeem M., 2014, Anatomical changes induced by NaCl stress in root and stem of Gazania harlequin L., Agricultural Commol/Lunications, 2(3): 8-14
Zapryanova N., and Atanassova B., 2009, Effects of salt stress on growth and flowering of ornamental annual species. In: XI Anniversary Scientific Conference, 120 Years of Academic Education in Biology, 45 Years Faculty of Biology, Institute of Ornamental Plants- Sofia, 1222, Negovan. pp. 177-179
http://dx.doi.org/10.1080/13102818.2009.10818394
Zidan I., Azaizeh H., and Neumann P. M., 1990. Does salinity reduce growth in maize root epidermal cells by inhibiting their capacity for cell wall acidification? Plant Physiology, 93: 7–11



. PDF(989KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Kekere Otitoloju
Related articles
. Strandline
. Salinity
. Anatomy
. Adaptation
Tools
. Email to a friend
. Post a comment