Estimation of Important Reproductive Parameters for Management of the Shoemaker Spinefoot Rabbitfish (Siganus sutor) in Southern Kenya 
 Author
Author  Correspondence author
Correspondence author
International Journal of Marine Science, 2012, Vol. 2, No. 4 doi: 10.5376/ijms.2012.02.0004
Received: 11 Sep., 2012 Accepted: 17 Sep., 2012 Published: 19 Sep., 2012
Agembe, 2012, Estimation of Important Reproductive Parameters for Management of the Shoemaker Spinefoot Rabbitfish (Siganus sutor) in Southern Kenya, International Journal of Marine Science, Vol.2, No.4 24-30 (doi: 10.5376/ijms.2012.02.0004)
The shoemaker spinefoot, Siganus sutor, is a member of the family Siganidae, commonly known as rabbitfish. Many demersal fish populations in the Kenyan waters are heavily exploited, and fishing effort may be above optimum levels for some species. This study was aimed at determining missing parameters on spawning periodicity and maturity of S. sutor for the Kenyan south coast area of Msambweni, complementing a suite of parallel studies on spawning aggregation based fisheries for this species. A total of 376 samples were collected over 12 months, including 85 that were collected over four lunar phases in November and December 2010. Monthly GSI showed two distinct peaks, one in June and July and one in November and January, suggested two distinct spawning periods within the year. The highest GSI was recorded in November while the lowest values occurred in April August and September. A lunar pattern in ovarian development stage was seen with the highest proportion of ripe ovaries (RI) observed in the full moon phase, whereas running ripe (RR) or hydrated ovaries dominated samples in the last quarter. Imminent spawning was confirmed by the presence of hydrated oocytes and post ovulatory follicles, observed primarily during the last quarter and to a lesser extent during full moon. GSI value Kruskal-Wallis test showed significant differences in GSI between moon phases (p< 0.05). In conclusion, histological analysis of ovaries points to synchronous development of oocytes with moon phases, suggest that their release takes place over short period of time after the full moon.
The Shoemaker spinefoot, Siganus sutor (Figure 1), is a member of the family Siganidae, commonly known as rabbitfish. The family is widely distributed in the Indian Ocean region (Lam, 1974; Randall, 1995) and is generally regarded as good food fishes in spite of their relatively small size. Woodland (1990) has described 13 siganid species that occur in the western Indian Ocean (WIO), including the endemic S. sutor. It occurs in coastal waters to a depth of at least 40 m and is important to artisanal and commercial fisheries. The species is caught with a variety of fishing gears (Woodland, 1984).
.png) Figure 1 Illustration of Siganus sutor (Source: Woodland, 1984) |
Many demersal fish populations in the Kenyan waters are heavily exploited, and fishing effort may be above optimum levels for some species (Kaunda-Arara et al., 2003; McClanahan and Omukoto, 2011). While S. sutor constitutes one of the most commercially important demersal fish resources in the country (de Souza, 1988), information on stock status is lacking and fisheries are not explicitly managed for this species. Knowledge of important reproductive parameters, such as the periodicity of manner of spawning can support management efforts aimed at protecting critical life history stages, including spawning aggregations. Spawning periodicity is also important for practical fisheries assessments i.e. estimating annual reproductive output and spawning potential ratios or spawning stock biomass per recruit (Sadovy et al., 2008).
Similarly, estimates of size at maturity can be used to improve gear-based management measures (e.g. mesh size regulations) or market-based measures, such as minimum size-at-sale. Spawning periodicity is also used to understand selectivity and assess levels of growth overfishing. Earlier studies on the reproductive biology of S. sutor had estimated size frequency distribution of the isolated oocytes, gonad maturity stages, breeding cycle, length-weight relationship and condition factor, but information on lunar periodicity of spawning and size at maturity was lacking (Ntiba and Jacccarini, 1988; de Souza 1988; Ntiba and Jacccarini, 1990).
The study was conducted at Msambweni in Kenya’s South Coast (Figure 2). Msambweni is a fishing settlement located about 70 km south of Mombasa town. It is experiencing the gradual influence of urbanization. The study area lies between latitude: 4°30'~4°35'S and longitude: 39°22~39°27' E in the South Coast region of Kenya (Muthiga and McClanahan, 1987). The study area is basically a low-lying coastal plain submerged complex (below 30 m contour) dominated by extensive cover of mangrove forests, intertidal areas covered with seagrass beds and coral reef in the shallow water lagoons. The past 30 years have seen the influx into this area of migrant fishermen who have introduced new fishing practices that have radically changed the fishery (McClanahan et al., 2005). The study focused on the main artisanal fisheries landing site Mkunguni/Msambweni This study was, therefore, aimed at determining missing parameters on spawning periodicity and maturity of S. sutor for the Kenyan south coast area of Msambweni, complementing a suite of parallel studies on spawning aggregation based fisheries for this species.
.png) Figure 2 Map of south coast of Kenya showing the fish landing site Msambweni monitored in the study (Source: Agembe et al., 2010) |
1 Results
A total of 376 samples were collected over 12 months, including 85 that were collected over four lunar phases in November and December 2010. Excluded from the present analysis were 177 individuals that were either immature, of undetermined sex. Monthly GSI showed two distinct peaks, one in June and July and one in November and January, suggested two distinct spawning periods within the year (Figure 3). The highest GSI was recorded in November while the lowest values occurred in April August and September. The monthly samples which were running ripe and spent ones are presented in (Figure 4) for comparison with the Monthly GSI graph (Figure 3) with highest ripe running fishes peaking November and January. Minor peaks recorded in June-July (Figure 4).
.png) Figure 3 Mean GSI (± standard error) by month over the period March 2009 ~ March 2010 for S. sutor females (n=199); GSI data were not collected in May and December 2009 |
.png) Figure 4 Gonad maturation for ripe running (RR) and spent fishes over the period March 2009 ~ February 2010 for S. sutor females (n = 107) |
Mean GSI by lunar phase (Figure 5) showed High GSI values occurring during the full moon. This period is not indicating that maximum spawning occurred that time. Whereas some were hydrated (high GSI), others could have just spawned.
.png) Figure 5 Mean GSI (± standard error) by lunar phase for November and December 2010 samples of S. sutor females (n = 99) |
A lunar pattern in ovarian development stage was seen with the highest proportion of ripe ovaries (RI) observed in the full moon phase, whereas running ripe (RR) or hydrated ovaries dominated samples in the first quarter (Figure 6). Resting (RE) ovaries predominated in the last quarter. Imminent spawning was confirmed by the presence of hydrated oocytes, which were observed primarily during the first quarter and to a lesser extent during full moon (Figure 6). Some ovaries had post-ovulatory follicles providing evidence of very recent spawning. These were observed in samples collected in the last week of the month of November 2010 GSI value Kruskal-Wallis test showed significant differences in GSI between moon phases (p<0.05). Nonetheless, hydrated ovaries were found in three of four lunar phases, while post-ovulatory follicles were observed in all lunar phases except the full moon phase, suggesting spawning in this species is protracted. Ovarian maturity stages are illustrated in Figure 7.
.png) Figure 6 Gonad maturation by lunar phase over a lunar cycle in November and December 2010 for S. sutor females (n = 85) |
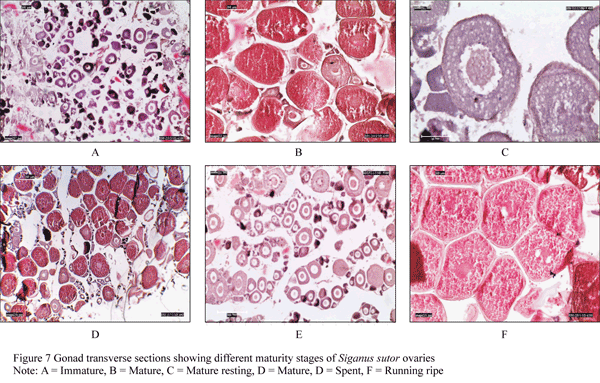 Figure 7 Gonad transverse sections showing different maturity stages of Siganus sutor ovaries |
The minimum size at first maturity, L50, the size at which 50% of females that were reproductively active, termed the effective maturity was estimated to be 28.2 cm TL (Figure 8).
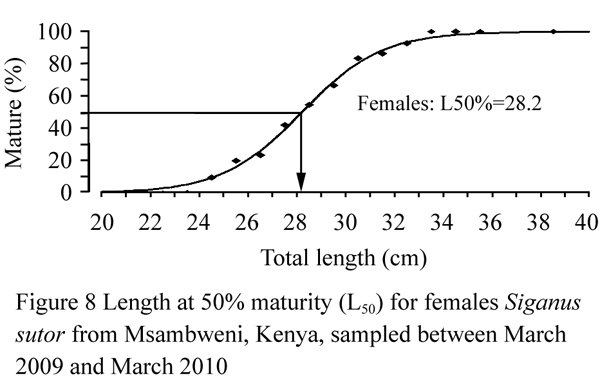 Figure 8 Length at 50% maturity (L50) for females Siganus sutor from Msambweni, Kenya, sampled between March 2009 and March 2010 |
2 Discussion
Trends in GSI revealed two distinct spawning peaks for S. sutor, a short peak in June-July and a more protracted peak from November to January/February. The month of October was observed as the likely period when the onset of ovary maturation occurs. These results concur with previous work on spawning seasonality of S. sutor by Ntiba and Jaccarini (1990), who also found two distinct peak spawning seasons for S. sutor in Kenyan waters (Table 1). In contrast, the current findings differ from those of de Souza (1988), which showed that S. sutor spawns throughout the year in Kenyan inshore waters. Tanzanian populations of S. sutor exhibit a less protracted spawning season than populations in Kenya (Table 1; Kamukuru, 2006). The difference in results may be explained by parameters like condition factor used in the other studies, which was assumed to relate to maturation schedules.
 Table 1 Macro- and microscopic criteria of female gonadal development (modified from de Souza, 1988; Ntiba and Jaccarini, 1990; Samoilys and Roelofs, 2000; West, 1990) |
In other studies bimodal spawning seasons in rabbitfishes as well as environmental cues such as temperature or photoperiod have been reported to possibly stimulate reproductive activity (Takemura et al., 2004). These findings suggest that a particular aspect of the lunar phase triggers synchronization in the final step of reproductive activity and that the lunar phase utilized is different among the rabbitfish species (Takemura et al., 2004).
The estimate of size at maturity in this study was considerably higher than that reported by earlier studies in the region (Ntiba and Jaccarini, 1988; Kamukuru, 2006). This partly relates to the parameter used with de Souza (1988), for example, reporting minimum size at maturity rather L50 maturity. In addition, prior studies did not use a logistic curve (see Table 2) (de Souza, 1988; Ntiba and Jaccarini, 1988; Kamukuru, 2006). Given that most ovaries sampled from fishes landed before midday were fully hydrated, and that hydration begins within 12 hours of ovulation (Sadovy, 1996), spawning in S. sutor may be occurring in the evening. S. sutor exhibited protracted spawning seasons corresponding to the NW monsoon with peaks in reproductive activity tending towards the inter-tropical monsoons, particularly October/November. Although the timing differs, a similar pattern has been demonstrated before for S. sutor in East Africa (Ntiba and Jaccarini, 1990), and a split spawning season may be common in areas affected by monsoons (Nzioka, 1979; Sadovy, 1996).
Table 2 Seasonality and minimum size at maturity of S. sutor in Kenya from previous studies and the present study |
In conclusion, histological analysis of ovaries points to synchronous development of oocytes with moon phases, which estimates that their release takes place over short period of time after the full moon. Or more specifically, spawning occurred during the last quarter moon phase. It is worthy to note that sampling only one month of spawning within one year is limited evidence for characterizing the spawning periodicity for the species across a year. It is possible for example, that the June–July spawning is a different population. Despite limited sample size, this study contributes to the state of knowledge about the spawning patterns of S. sutor in the Msambweni area, which can guide future management measures for a species that is subjected to spawning aggregation fishing. The management should consider temporal catch and sales ban inclusive of November-January and June-July. This study provides further impetus for directing research to environmental cues in the future.
3 Materials and Methods
To identify seasonal and lunar reproductive periodicity and size at sexual maturity for S. sutor in Msambweni, southern Kenya, artisanal fishery catches were sampled during 11 of 13 months (March 2009~March 2010, excluding May and December 2009) at a coastal landing site. Sampling was stratified, based to reported reproductive (November-March: Ntiba and Jaccarini, 1988) and non-reproductive periods, with 5 days sampled per month during the spawning season and 3 days per month for the remainder of the year. Sampling trips focused on new moon periods, but included other lunar phases during one lunar cycle from November to December 2010. Females were sampled preferentially as ovarian development is more reliable for detailing spawning periodicity (West, 1990).
Fish were sampled for length (TL (from snout to tip of longest lobe of caudal fin), nearest mm) and wet weight (nearest 0.1 g), and gonads removed and weighed (nearest 0.01 g). To determine lunar periodicity in spawning, a sub-set of samples collected weekly over one complete lunar cycle were evaluated microscopically with gonad maturity stage assessed using a developmental scale ranging from immature to spent (Table 1). To document lunar periodicity and confirm seasonal periodicity, gonado-somatic index (GSI) for females was estimated using the following formula:
GSI = weight of ovary (g)/weight of fish (g) ï¼ weight of ovary (g) × 100.
Sampled gonads were preserved in a 10% formalin solution, buffered with acetic acid and calcium chloride (FAACC) for histological preparations. Microscopic staging was based on standard techniques (Ntiba and Jaccarini, 1990; West, 1990; Samoilys and Roelofs, 2000). An automatic tissue processor (Leica TP 1020, Wetzlar, Germany) was used to process haematoxylin/eosin-stained tissue samples which were then sectioned at 5~7 µm. Reading of slides was done by two observers using Table 1 criteria.
The minimum size at first maturity, L50, the size at which 50% of females that were reproductively active during the main spawning periods, was estimated by fitting a logistic curve, using the least square method, to the proportion of macroscopically mature individuals by size class (see Kolding and Skaalevik, 2010). This method, termed as effective maturity recognises that, despite maturity, not all females are reproductively active in the spawning season (Pears et al., 2006), and also minimises errors in assigning immature fish as inactive females (Samoilys and Roelofs, 2000). Gonads that were undetermined in terms of maturity stage were excluded from this assessment.
Authors Contributions
SA completed data analysis, essayed and provided techniques for data analysis, designed the research, guided the process, prepared and modified this paper.
Acknowledgements
The author is grateful to the Director Kenya Marine & Fisheries Research Institute (KMFRI) for laboratory space and to Boaz Orembo and James Gonda who assisted in data collection and histological laboratory analyses at KMFRI. Dr. Melita Samoilys is thanked for providing guidance and advice both in the field and in the laboratory. Thanks to Jan Robinson for providing good suggestions during preparation of this chapter. Much thanks to Dr. Kevin Rhodes for useful comments when he reviewed this chapter. The author is grateful to the anonymous reviewer for useful comments. This project was funded by Western Indian Ocean Marine Science Association (WIOMSA) through Marine and Coastal Science for Management (MASMA) programme.
References
Agembe S., Mlewa C.M., Kaunda-Arara B., 2010, Catch composition, abundance and length-weight relationships of groupers (Pisces: Serranidae) from inshore waters of Kenya, Western Indian Ocean J. Mar. Sci., 9 (1): 239 - 248
De Souza T., 1988, Reproduction, length-weight relationship and condition factor in Siganus sutor (Valenciennes, 1835) (Pisces: Siganidae) from the Kenyan waters of the western Indian Ocean, Kenya Journal of Science Series B, 9: 89-101
Kamukuru A.T., 2006, Reproductive biology of white spotted rabbitfish, Siganus sutor (Pisces: Siganidae) from basket trap fishery in Dar es Salam marine reserve systems, Tanzania, WIOMSA MARG I/2006-01, pp. 31
Kaunda-Arara B., Rose G.A., Muchiri M.S., and Kaka R., 2003, Long-term trends in coral reef fish yields and exploitation rates of commercial species from coastal Kenya, Western Indian Ocean J Mar Sci, 2:105-116
Kolding J., and Skålevik A., eds., 2010, Pasgear II Version 2.4. University of Bregen. Bregen, Norway
Lam T.J., 1974, Siganids: Their biology and mariculture potential, Aquaculture, 3:325-354
http://dx.doi.org/10.1016/0044-8486(74)90001-5
McClanahan T.R., Mwanguni S., and Muthiga N.A., 2005, Management of the Kenyan Coast, Ocean and Coastal Management, 48: 11-22
http://dx.doi.org/10.1016/j.ocecoaman.2005.03.005
McClanahan T.R., and Omukoto J.O., 2011, Comparison of Modern and Historical Fish Catches (AD 750–1400) to Inform Goals for Marine Protected Areas and Sustainable Fisheries, Conserv Biol 25(5): 945-955
http://dx.doi.org/10.1111/j.1523-1739.2011.01694.x PMid:21676028
Muthiga N., and McClanahan T.R., 1987, Population changes of sea urchin Echinometra mathae on an exploited fishing reef, African journal of ecology, 25: 1-8
http://dx.doi.org/10.1111/j.1365-2028.1987.tb01085.x
Ntiba M.J., and Jaccarini V., 1988, Age and growth parameters of Siganus sutor in Kenyan marine inshore water, derived from number of otolith microbands and fish length, Journal of Fish Biology, 33: 465-470
http://dx.doi.org/10.1111/j.1095-8649.1988.tb05487.x
Ntiba M.J., and Jaccarini V., 1990, Gonad maturation and spawning times of Siganus sutor off the Kenya coast: evidence for definite spawning seasons in a tropical fish, J Fish Biol, 37: 315-325
http://dx.doi.org/10.1111/j.1095-8649.1990.tb05862.x
Nzioka R.M., 1979, Observation on the spawning seasons of East African reef fishes, J. Fish Biol., 14: 329-342
http://dx.doi.org/10.1111/j.1095-8649.1979.tb03528.x
Pears R.J., Choat J.H., Mapstone B.D., and Begg G.A., 2006, Demography of a large grouper Epinephelus fuscoguttatus, from Australia’s Great Barrier Reef: Implications for fishery management, Mar Ecol Prog Ser, 307: 259-272
http://dx.doi.org/10.3354/meps307259
Randall J.E., ed., 1995, Coastal fishes of Oman. University of Hawaii Press, Honolulu, HI, pp.439
Sadovy de Mitcheson Y., Cornish A., Domeier M., Colin P.L., Russell M., and Lindeman K.C., 2008, A global baseline for spawning aggregations of reef fishes, Conserv Biol, 22:1233-1244
http://dx.doi.org/10.1111/j.1523-1739.2008.01020.x PMid:18717693
Sadovy Y., 1996, Reproduction of reef fishery species. In: Reef Fisheries. Polunin NVC, Roberts CM (Eds), Chapman and Hall, London, pp. 15-59
Samoilys M.A., and Roelofs A., eds., 2000, Defining the reproductive biology of a large serranid Plectropomus leopardus. CRC Reef Research Center Technical Manual No. 31, CRC Research Centre, Townsville, Australia, pp.36
Takemura A., Rahman M.S., Nakamura S., Park Y.J., and Takano K., 2004, Lunar cycles and reproductive activity in reef fishes with particular attention to rabbitfishes, Fish and Fisheries, 5: 317-328
http://dx.doi.org/10.1111/j.1467-2679.2004.00164.x
West G., 1990, Methods of assessing ovarian development in fishes: A review, Aust J Mar Freshwater Res, 41:199-222
http://dx.doi.org/10.1071/MF9900199
Woodland D.J., ed., 1984, Aid to the identification of fish family Siganidae. In: FAO Species Identification Sheets for Fishery Purposes: Western Indian Ocean, Fishing Area 51, Vol 4. Fisher W, Bianchi G (Eds.), FAO, Rome
Woodland D.J., 1990, Revision of the fish family Siganidae with descriptions of two new species and comments on distribution and biology, Indo-Pacific Fishes, 19: 136



. PDF(1074KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Simon Agembe
Related articles
. Shoemaker spinefoot rabbitfish
. Siganus sutor
. South coast Kenya
. Reproductive parameters
. Histology
. Lunar phase
Tools
. Email to a friend
. Post a comment